Resveratroli on monissa kasveissa ja mm. punaviinissä esiintyvä fenoliyhdiste, jolla uskotaan olevan verrattomia terveysvaikutuksia. Resveratroli voi ehkäistä pahanlaatuisten syöpäsolujen lisääntymistä, torjua virustauteja, hidastaa vanhenemista ja hillitä elimistöä rasittavaa hiljaista tulehdusta. Resveratroli saattaa myös suojaata hermosoluja.
Mitä resveratroli on?
Resveratroli (trans-3,5,4-trihydroksistilbeeni) on fytoaleksiini, eli kasvien tuottama entsyymi, joka puolustaa kasveja bakteeri- ja sienihyökkäyksiltä.
Fytoaleksiinit kuuluvat kasveja patogeeneiltä suojaaviin fytokemikaaleihin, kuten terpenoidit, fenoliyhdisteet, glykosteroidit ja alkaloidit. Fytokemikaalit ovat osa kasvien luonnollista puolustusjärjestelmää.
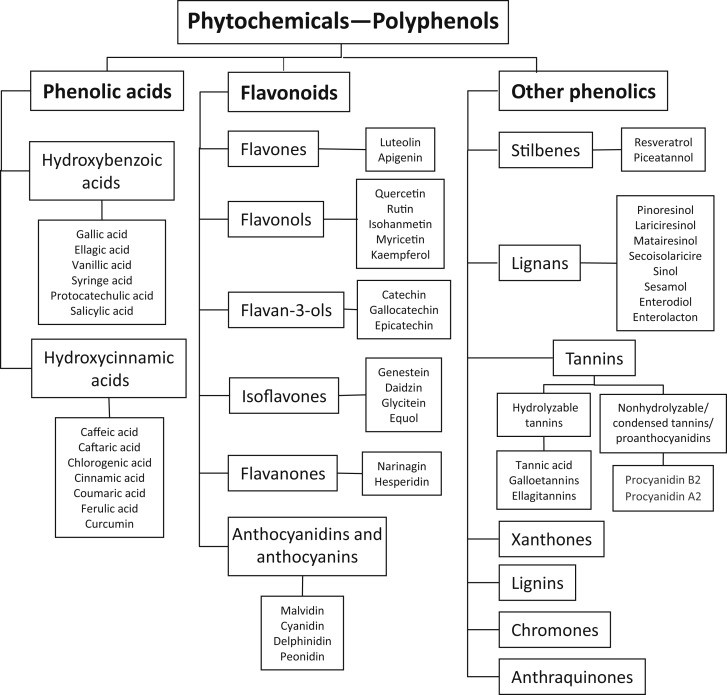
Fytokemikaalit ja flavonoidit
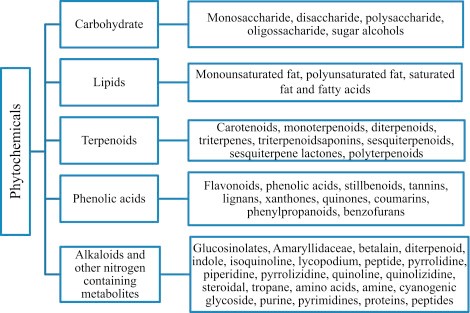
”Generally, phytochemicals have been classified into six major categories based on their chemical structures and characteristics. These categories include carbohydrate, lipids, phenolics, terpenoids and alkaloids, and other nitrogen-containing compounds (Figure 1; Harborne and Baxter, 1993; Campos-Vega and Oomah, 2013).”
Kuvan lähde: ScienceDirect
Fytokemikaalit ovat kasveissa esiintyviä biologisesti aktiivisia kemiallisia yhdisteitä. Flavonoidit ovat fytokemikaaleihin kuuluvia kasveissa muodostuvia fenoliyhdisteitä. Ne antavat kasveille, niiden kukille, hedelmille ja siemenille värin ja suojaavat kasveja ultraviolettisäteilyltä.
Ainakin osa flavonoideista on antioksidantteja, eli ne suojelevat elimistöä vapaiden happiradikaalien vaikutukselta. Erilaisia flavonoideja on löydetty joitakin tuhansia ja niitä löydetään koko ajan enemmän. Ennen 1950-lukua flavonoideja kutsuttiin parin vuosikymmenen ajan P-vitamiineiksi.
Resveratroli, jota esiintyy runsaasti mm. viinirypäleissä, viininlehdissä, mustikoissa, maapähkinöissä ja karpaloissa, on tällainen oksidatiivista stressiä hillitsevä antioksidantti. Sillä on myös antimikrobisia eli mikrobeja tuhoavia ominaisuuksia.
Punaviini sisältää resveratrolia. Aika ajoin julkaistaan (viiniteollisuuden lobbaamia?) tutkimuksia, joiden mukaan lasillinen punaviiniä päivässä suojelee sydäntä ja hillitsee elimistön matala-asteista tulehdusta. Vallitseva ravintotieteellinen näkemys taitaa kuitenkin olla, että punaviinin terveyshaitat ylittävät sen potentiaaliset terveyshyödyt.
Esimerkiksi ranskalaisten alhaisen sydän- ja verisuonitautiesiintyvyyden on väitetty johtuvan runsaasti resveratrolia sisältävän punaviinin kulutuksesta. Toisaalta ranskalaista paradoksia on selitetty myös luonnollisten rasvojen runsaalla saannilla.
Resvetatroli ja syöpä
Useissa tutkimuksissa on havaittu, että resveratroli voi laskea syöpään sairastumisen riskiä. Hiirikokeissa terveyttä edistäväksi ja syövältä suojaavaksi annokseksi osoitettiin määrä, jonka ihminen saisi kuitenkin vasta neljälläsadalla punaviinilasillisesta päivässä.
Onko resveratroli syövältä suojaava tai syövän parantava ihmeyhdiste? Resveratrolin kohtuullinen saanti voi ylläpitää kehon hyvinvointia antioksidanttina ja sirt1-geenin aktivaation kautta.
Eräs tuore tutkimus havainnollisti kuinka resveratroli voi estää useimmissa rintasyövissä esiintyvän mutatoituneen proteiinin lisääntymisen. Tämän vaikutuksen uskotaan edistävän elimistön kykyä taistella syöpäsolujen lisääntymistä vastaan.
Resveratrolin terveysvaikutukset
Resveratrolia markkinoidaan monenlaisilla terveyshyödyillä. Sen uskotaan hidastavan aivojen vanhenemista, ehkäisevän aivojen inflammaatiota dementiaa sairastavilla ja taistelevan syöpää vastaan. Sitä myös kaupataan tärkeänä lisäravinteena.
Korrelaatio resveratrolin ja terveyshyötyjen välillä on olemassa. Sen sijaan kausaliteetti on hankalampi osoittaa. Ainakin joissain tapauksissa resveratrolin terveysvaikutuksia on tutkimuksissa korostettu kaupallisten syiden vuoksi. Antioksidanttina se mahdollisesti suojaa hapetusstressiltä ja inflammaatiolta, jotka ovat taustatekijöitä useimmissa yleisissä sairauksissa.
p53-proteiini
Resveratrolin syöpää ehkäisevät ominaisuudet liittyvät proteiiniin p53. Tämän proteiinin mutaatioiden aggregoituminen assosioituu yli 50 prosenttiin syöpäkasvaimista.
Brasilialaistutkijat ovat osoittaneet, että resveratroli estää aktiivisesti mutatoituneiden proteiinien aggregoitumista ja ehkäisee syöpäsolujen lisääntymistä ja leviämistä muualle kehoon.
p53-proteiini on terveessä elimistössä eräänlainen ”genomin vartija”, sillä se tuhoaa luontaisesti pahanlaatuisia syöpäsoluja ja suojelee terveitä soluja.
Mutatoitunut p53-proteiini on kuitenkin haitallinen elimistölle. Se muodostaa kasvavia amyloidiplakkeja. Amyloidit ovat proteiinifragmentteja, joita muodostuu aineenvaihdunnan seurauksena.
Eräissä aineenvaihdunnan häiriöissä amyloidit kertyvät elimistöön amyloidiplakeiksi, joka aiheuttaa amyloidoosia. Amyloidit assosioituvat mm. Parkinsonin tautiin, nivelreumaan ja Huntingtonin tautiin. Alzheimerin taudissa beeta-amyloideja kertyy aivoihin, jossa ne aiheuttavat muistista vastaavien aivosolujen kuolemaa.
Amyloidien ja erityisesti p53-proteiinien kasautumisen plakeiksi uskotaan assosioituvan myös eräiden syöpien kehittymiseen.
Verrattain uudessa tutkimuksessa da Costa tutkimusryhmineen sovelsi fluoresenssia spektroskopiaa selvittääkseen in vitro (koeputkessa) kuinka resveratroli vaikuttaa mutatoituneisiin p53-proteiineihin.
Lisäksi da Costan tutkijaryhmä toteutti immunofluoresenssin kolokalisaation analyysejä selvittääkseen resveratrolin vaikutuksen rintasyöpäsolujen solulinjoihin, joissa esiintyi p53-proteiinin mutaatioita sekä rintasyöpäsoluihin, joissa esiintyi normaaleja p53-proteiineja. Rintasyöpäsoluja istutettiin hiirille, joilla testattiin resveratrolin vaikutusta pahanlaatuisiin kasvaimiin.
Laboratoriokokeet osoittivat, että resveratroli esti p53-proteiinien kasaantumisen sekä in vitro ihmisten rintasyöpäsoluihin ja hiirien pahanlaatuisiin kasvaimiin.
Lisäksi resveratroli vähensi da Costan tutkimusryhmän mukaan merkittävästi pahanlaatuisten solujen lisääntymistä ja leviämistä.
Nyt saadut tutkimustulokset auttavat da Costan mukaan kehittämään lääkettä, joka voi ehkäistä mutatoituneiden p53-proteiinien aggregoitumista amyloidiplakeiksi.
”This study provides evidence that resveratrol directly modulates p53 and enhances our understanding of the mechanisms involved in p53 aggregation as a therapeutic strategy for cancer treatment. Our data indicate that resveratrol is a highly promising lead compound targeted against mutant p53 aggregation.”
”Resveratrol is a natural phytoestrogen with neuroprotective properties. Polyphenolic compounds including resveratrol exert in vitro antioxidant, anti-inflammatory, and antiamyloid effects. Resveratrol and its derivative pterostilbene are able to cross the blood-brain barrier and to influence brain activity. The present short review summarizes the available evidence regarding the effects of these polyphenols on pathology and cognition in animal models and human subjects with dementia. Numerous investigations in cellular and mammalian models have associated resveratrol and pterostilbene with protection against dementia syndromes such as Alzheimer’s disease (AD) and vascular dementia. The neuroprotective activity of resveratrol and pterostilbene demonstrated in in vitro and in vivo studies suggests a promising role for these compounds in the prevention and treatment of dementia. In comparison to resveratrol, pterostilbene appears to be more effective in combatting brain changes associated with aging. This may be attributed to the more lipophilic nature of pterostilbene with its two methoxyl groups compared with the two hydroxyl groups of resveratrol. The findings of available intervention trials of resveratrol in individuals with mild cognitive impairment or AD do not provide evidence of neuroprotective or therapeutic effects. Future clinical trials should be conducted with long-term exposure to preparations of resveratrol and pterostilbene with high bioavailability.” Lähde: PubMed
Hiljattain joukko amerikkalaisia professoreita määritteli sokerin terveydelle haitallisimmaksi ravintoaineeksi. Sokerin terveydelliset vaikutukset palautuvat erityisesti glukoosin ja fruktoosin aineenvaihduntaan. Kuinka glykolyysi ja fruktolyysi eroavat toisistaan?
Sokereiden aineenvaihdunta
Ravinto sisältää useita sokereita erilaisina molekyylirakenteina. Niiden pilkkominen imeytyvään muotoon alkaa jo suussa. Yleisimmät ravinnosta saatavat sokerit ovat, glukoosi, fruktoosi, laktoosi, galaktoosi ja maltoosi.
Kasvien sisältämä tärkkelys on sokerin varastomuoto. Tärkkelys on monimutkainen glukoosiyksiköistä muodostuva haarautumaton amyloosi tai haarautunut amylopektiini. Se on polysakkaridi, jossa glukoosimonomeerit ovat liittyneet toisiinsa. Kasvit valmistavat tärkkelystä fotosynteesissä. Erityisen tärkkelyspitoisia ravintoaineita ovat esimerkiksi perunat, maissi ja riisi.
Solujen energiantuotannon kannalta glukoosi ja fruktoosi ovat tärkeimpiä sokereita. Niilläkin on omat aineenvaihduntapolkunsa.
Sokereita kuljettavat molekyylit
Glukoosimolekyylejä kuljettavat ohutsuolesta verenkiertoon insuliinin säätelemät GLUT1- ja GLUT4-molekyylit. Fruktoosi ei aktivoi insuliinintuotantoa ja sitä kuljettavat GLUT5-molekyylit.
Käytännössä GLUT1-molekyyli kuljettaa glukoosimolekyylin ohutsuolen endoteelisoluun ja GLUT4-molekyyli kuljettaa glukoosimolekyylin endoteelisolusta verenkiertoon.
Ruoansulatuskanavassa sokereita pilkkoo joukko entsyymejä, joista tärkkelystä maltoosiksi pilkkova amylaasi on ehkä tutuin. Amylaasi aloittaa tärkkelyksen pilkkomisen jo suussa, johon amylaasia erittyy sylkirauhasista. Sokereiden pilkkominen imeytyvään muotoon jatkuu vatsassa ja suolistossa. Haima erittää amylaasia ohutsuoleen.
Maltoosi on kahdesta glukoosimolekyylistä muodostuva disakkaridi. Ruoansulatuskanavassa maltoosia pilkkoo suolinesteen maltaasi-entsyymi. Laktoosi eli maitosokeri koostuu glukoosista ja galaktoosista. Laktoosia pilkkoo laktaasi. Sakkaroosi muodostuu fruktoosista ja glukoosista. Sakkaroosia pilkkoo sakkaraasi.
Ruoansulatuskanavassa sokerit pilkotaan yksinkertaisiksi sokerimolekyyleiksi, jotka kulkeutuvat ohutsuolesta verenkertoon sokereille spesifien kuljetusmolekyylien kuljettamina. Sokereiden aineenvaihduntareitit poikkeavat toisistaan.
Esimerkiksi: Aldoosi-1-epimeraasi (GALM) muuttaa β-D-galaktoosin ensin α-D-galaktoosiksi. Tämä muutetaan edelleen UDP-glukoosiksi kolmen pääasiallisen entsyymin avulla (GALK, GALT ja GALE). UDP-glukoosi (uracil-diphosphate glucose) on nukleotidin, eli nukleiinihappojen (DNA, RNA) rakenneyksikön sokeriosa. Nukleotidiin kuuluu kolme osaa, emäs, sokeriosa ja fosfaatti. Galaktoosi vaikuttaa siis nukleiinihappojen aineenvaihduntaan ja rakenteeseen.
Pöytäsokeri on fruktoosia ja glukoosia
Tavallinen pöytäsokeri on ruokosokeria eli sakkaroosia, joka muodostuu yhtäläisestä määrästä glukoosia ja fruktoosia. Sakkaroosi on disakkaridi, jossa kaksi erilaista sokerimolekyyliä on tiukasti sitoutunut toisiinsa.
Glukoosi (rypälesokeri) on elimistön tärkein energianlähde. Glukoosi on yhdestä sokerimolekyylistä muodostuva monosakkaridi. Myös glukoosia selvästi makeampi fruktoosi (hedelmäsokeri) on yhdestä sokerimolekyylistä muodostuva monosakkaridi.
Kasveissa sakkaroosi on yleinen. Sitä on paljon esimerkiksi sokeriruo’ossa, sokerijuurikkaassa, ananaksessa, maississa ja porkkanassa.
Ruoansulatus pilkkoo hiilihydraateista ja tärkkelyksestä yksinkertaisia ohutsuolesta verenkiertoon imeytyviä sokerimolekyylejä. Glukoosi ja fruktoosi ovat yleisimmät ravinnosta saatavat sokerit, mutta ne eivät toimi elimistössä aivan samalla tavalla. Näiden sokereiden reaktioketjut eroavat aineenvaihdunnan kannalta merkittävästi toisistaan.
Fruktoosi ja fruktolyysi
Fruktoosi
Fruktoosi on yhdestä fruktoosimolekyylistä muodostuva monosakkaridi, jota esiintyy luonnostaan hedelmissä, marjoissa ja vihanneksissa joko vapaina fruktoosimolekyyleinä, kahdesta sokerimolekyylistä muodostuvina disakkarideina (kuten ruokosokeri) tai fruktoosin polymeerina (inuliini).
Inuliini on fruktoosista muodostunut varastopolysakkaridi, jossa on 30-50 fruktoosimolekyyliä. Inuliinia esiintyy mm. maa-artisokassa, sipulissa, parsassa, banaanissa, rukiissa, vehnässä, mustajuuressa, ohrassa sekä asterikasvien, kuten daalian, voikukan ja sikurin juurissa ja juurimukuloissa. Inuliinia käytetään elintarviketeollisuudessa makeutusaineena. Se sisältää noin kolmanneksen vastaavan sokerimäärän energiasta.
Teollinen fruktoosi
Fruktoosi on makeampaa kuin glukoosi, joten sitä käytetään yleisesti makeutusaineena. Tavallinen kidesokeri sisältää 50:50 suhteessa glukoosia ja fruktoosia. Fruktoosia saa runsaasti mm. hedelmistä ja marjoista. Teollisesti valmistettua fruktoosia ja fruktoosisiirappia käytetään yhä useammissa elintarvikkeissa.
Edullinen nestemäinen fruktoosisiirappi on syrjäyttämässä perinteisen sakkaroosin yleisimpänä makeutusaineena. Amerikkalaisessa ruokavaliossa päivittäisestä energiansaannista jopa 10 % saadaan fruktoosista. Jos suomalaisen päivittäisestä energiansaannista 10% on peräisin lisätystä sokerista, silloin fruktoosin osuus energiansaannista voi olla jopa 5 %.
Teollinen fruktoosi korreloi vahvasti monien kardiometabolisten sairauksien kanssa. Tätä selittää todennäköisesti fruktoosin glukoosista poikkeava aineenvaihdunta. Erityisen ongelmallista on fruktoosisiirapista nopeasti elimistöön imeytyvät vapaat fruktoosimolekyylit, jotka voivat horjuttaa sokeriaineenvaihdunnan toimintaa, rasittaa maksaa ja altistaa esimerkiksi alkoholista riippumattomalle rasvamaksalle.
Huomio: Korrelaatio fruktoosisiirapin ja kardiometabolisten sairauksien välillä on vahva, mutta kausaliteettia ei välttämättä ole osoitettu.
Fruktolyysi
Fruktoosin aineenvaihdunta tapahtuu pääasiassa maksassa. Maksan lisäksi fruktolyysiä tapahtuu jonkin verran ohutsuolessa, luurankolihaksissa, kiveksissä, rasvakudoksessa ja aivoissa.
Noin prosentti syödystä fruktoosista muutetaan maksassa suoraan plasman triglyserideiksi. 29-54% fruktoosista syntetisoidaan maksassa glukoosiksi. Neljännes fruktoosista muutetaan laktaatiksi eli maitohapon suoloiksi tai estereiksi. 15-18% fruktoosista syntetisoiduista glykoosimolekyyleistä varastoidaan glykogeenien polymerisaatiossa polysakkarideina glykogeeneiksi.
Glykogeenit ovat tuhansista glukoosimolekyyleistä muodostavia pitkäketjuisia ja pitkähaaraisia polysakkarideja.
Glykogeenien polymerisaatio
Glykogeenin polymerisaation lähtöaineina toimivat sokeri-nukleotidit, joissa sokerimonomeeri on aktivoitu kiinnittämällä siihen nukleotidi. Glykogeenin polymerisaatiossa vaikuttavat glykogeenisyntaasi, joka liittää glukoosimolekyylejä pitkään ketjuun sekä entsyymi, joka tekee glykogeeniin haarakohtia. Koska glykogeenisyntaasi voi vain lisätä glukoosimolekyylejä valmiiseen ketjuun, tarvitaan oma entsyymi aloittamaan glykogeenin valmistus (glykogeniini). Lähde: Solunetti
Kehon solut voivat käyttää fruktolyysin syntetisoimaa glukoosia ja laktaattia energianlähteenä solujen glykolyysissä. Aineenvaihdunta voi myös purkaa maksaan ja lihassoluihin varastoituneita fruktoosista valmistettuja glykogeenejä glukagonin stimuloimana glykogenolyysissä, jolloin vereen vapautuu glukoosia. Tämä mekanismi turvaa solujen energiansaannin ruokailujen välillä sekä raskaissa fyysisissä suorituksissa.
Alhainen verensokeri aktivoi haiman erittämään glukagonia, joka purkaa maksan ja lihasten glykogeenejä glukoosiksi. Glukagoni aktivoi myös maksassa ja munuaisissa tapahtuvaa glukoneogeneesiä, ketogeneesiä sekä rasvojen β-oksidaatiota. Matala verensokeri aktivoi rasvan käyttämistä energianlähteenä.
Glykolyysi
Kaikki solut saavat energiaa glukoosista. Solu saa energiantuotantoon tarvittavan glukoosin joko solun ulkopuolelta, josta se kulkeutuu soluun osmoottisesti eli suuremmasta pitoisuudesta pienempään, tai purkamalla solun sisäisen glykogeenin glukoosimolekyyleiksi.
Glykolyysi on solulimassa tapahtuva monimutkainen reaktiosarja, jossa glukoosimolekyylit hajotetaan kahdeksi palorypälehapon anionimuodoksi eli pyruvaatiksi.
Yhdestä glukoosimolekyylistä saadaan kahden pyruvaattimolekyylin lisäksi kaksi ATP-molekyyliä ja kaksi NADH-molekyyliä. Solujen anaerobinen energiansaanti perustuu glykolyysiin.
Soluhengitys
Jos solussa on mitokondrioita ja happea, energiantuotanto jatkuu sitruunahappokierrossa (Krebsin sykli). Eräiden entsyymien avustuksella pyruvaateista saadaan mitokondrioissa tapahtuvassa oksidatiivisessa dekarboksylaatiossa kaksi asetyylikoentsyymi-A:ta.
Rasvahapot hajoavat energiaksi pääasiassa hapettumalla β-oksidaatiossa siten, että rasvahappoketjusta irtoaa kahden hiilen asetyyliryhmiä, jotka ovat kiinnittyneenä reaktioon osallistuvaan koentsyymi-A:han.
Asetyylikoentsyymi-A eli aktiivinen etikkahappo on kaikille ravintoaineille yhteinen välituote solun valmistaessa energiaa. Asetyylikoentsyymi-A:ta saadaan monosakkarideista, triglyserideista sekä aminohapoista erilaisten reaktiovaiheiden kautta. Asetyylikoentsyymi-A:n asetyyliryhmän hiilet hapettuvat hiilidioksidiksi Krebsin syklissä ja vedyt siirtyvät erityisten koentsyymien avulla elektroninsiirtoketjuun. Näissä reaktioissa syntyy energiaa, joka varastoidaan fosfaattiyhdisteisiin, esimerkiksi ATP:ksi. Lähde: Wikipedia
Anaerobinen ja aerobinen glykolyysi
Soluissa, joissa ei ole mitokondrioita (esim. veren punasolut) tai riittävästi happea, pyruvaatti pelkistyy maitohapoksi. Maitohappoon pelkistyvää glykolyysiä kutsutaan anaerobiseksi glykolyysiksi ja asetyylikoentsyymi-A:han päättyvää glykolyysiä aerobiseksi glykolyysiksi.
Asetyylikoentsyymi-A:han päättyvän reaktiossa glukoosimolekyylistä jää jäännöstuotteena hiilidioksidia ja vettä, jotka poistuvat kehosta ihon ja hengityksen kautta.
Laktaatin muodostuminen ei tuota energiaa, mutta se on välttämätöntä, jotta saadaan pelkistyneet NADH-molekyylit takaisin hapettuneeseen NAD+-muotoon, jota tarvitaan glykolyysissä.
Glykolyysin vaiheet (Lähde: Wikipedia – Glykolyysi)
Glukoosiin liitetään fosfaattiryhmä ATP:ltä heksokinaasin tai glukokinaasin avulla, jolloin syntyy glukoosi-6-fosfaattia. Heksokinaasia inhiboi glukoosi-6-fosfaatti ja glukokinaasia fruktoosi-6-fosfaatti. Lisäksi insuliini aktivoi glukokinaasin transkriptiota tumassa.
Glukoosi-6-fosfaatti muutetaan fosfoheksoosi-isomeraasin avulla fruktoosi-6-fosfaatiksi.
Fruktoosi-6-fosfaatti fosforyloidaan ATP:n avulla fruktoosi-1,6-bisfosfaatiksi. Tätä reaktiota katalysoi fosfofruktokinaasi-1 eli PFK-1. PFK-1:tä inhiboi ATP, sitraatti ja H+. Aktivoivia molekyylejä ovat puolestaan AMP ja fruktoosi-2,6-bisfosfaatti.
Fruktoosi-1,6-bisfosfaatti muutetaan aldolaasi A:n:n avulla dihydroksiasetonifosfaatiksi ja glyseraldehydi-3-fosfaatiksi.
Dihydroksiasetonifosfaatti muutetaan trioosifosfaatti-isomeraasin avulla glyseraldehydi-3-fosfaatiksi.
Neljännen ja viidennen vaiheen reaktioista saaduista glyseraldehydi-3-fosfaateista muodostetaan glyseraldehydi-3-rosfaattidehydrogenaasin avulla 1,3-bisfosfoglyseraattia. Samalla NAD+ pelkistyy NADH:ksi.
1,3-bisfosfoglyseraatti defosforyloidaan fosfoglyseraattikinaasin avulla 3-fosfoglyseraatiksi. Samalla ADP fosforyloituu ATP:ksi.
3-fosfoglyseraatti muutetaan fosfoglyseraattimutaasin avulla 2-fosfoglyseraatiksi.
2-fosfoglyseraatti muutetaan enolaasin avulla fosfoenolipyruvaatiksi.
Fosfoenolipyruvaatti defosforyloidaan pyruvaattikinaasin avulla pyruvaatiksi. Samalla ADP fosforyloituu ATP:ksi. Pyruvaattikinaasia inhiboivat ATP, alaniini ja glukagoni. Aktivoiva molekyyli on puolestaan glykolyysin kolmannessa vaiheessa muodostuva fruktoosi-1,6-bisfosfaatti.
Glukoosin ja fruktoosin aineenvaihduntaan osallistuu samoja entsyymejä ja solujen rakenteita sekä niitä yhdistäviä reaktioketjuja, mutta monista yhtäläisyyksistä huolimatta niiden aineenvaihdunta eroaa toisistaan merkittävällä tavalla.
Fruktoosi ei stimuloi insuliinin eritystä
Fruktoosi ei stimuloi insuliinin eritystä samalla tavalla kuin glukoosi, eikä sen pääsy soluihin ole insuliinista riippuvainen. Glukoosin aineenvaihdunta puolestaan tarvitsee insuliinia.
Insuliinin tuotannon loppuminen tyypin 1 diabeteksessa sekä solujen kasvanut insuliiniresistenssi tyypin 2 diabeteksessa aiheuttavat sen, että solut eivät saa energiantuotannossa tärkeää glukoosia, vaan glukoosimolekyylit jäävät verenkiertoon, jossa ne vaurioittavat verisuonia ja sisäelimiä.
Veren glukoosipitoisuus stimuloi haiman insuliinineritystä. Insuliinin säätelemät kuljetusmolekyylit (GLUT1 ja GLUT4) kuljettavat glukoosin soluihin. Fruktoosin kuljetusmolekyyli on GLUT5.
Haiman erittämä insuliini kiinnittyy solujen insuliinireseptoreihin, mikä käynnistää soluissa toisioviestintäjärjestelmän. Se houkuttelee solun sisällä olevan solukalvon läpäisevän glukoosinsiirtokanavan solukalvolle. Glukoosi pääsee tämän avulla solun sisälle, jossa glukoosimolekyyli osallistuu energiaa tuottavaan glykolyysiin.
Sokereiden rakenne
Monosakkaridit, kuten fruktoosi ja glukoosi voivat olla joko avoketjuisia tai renkaita, mutta elimistössä monosakkaridien vallitsevana muotona on rengasmainen rakenne.
Rengasmaisessa rakenteessa anomeerihiileksi kutsutaan hiiltä, joka on lähimpänä karbonyyliryhmää muodostavaa hiiltä. Karbonyyliryhmästä aloitetaan monosakkaridien hiilten numerointi. Hiiliketjun pituus monosakkaridissa voi olla kolme tai suurempi. Tärkeimpiä elimistössä esiintyviä monosakkarideja ovat heksoosit, joissa on kuuden hiiliatomin ketju.
Trioosi: kolme hiiliatomia
Totroosi: neljä hiiliatomia
Pentoosi: viisi hiiliatomia
Heksoosi: kuusi hiiliatomia
Heptoosi: seitsemän hiiliatomia
Karbonyyliryhmä koostuu toisiinsa kaksoissidoksella kiinnittyneistä hiili- ja happiatomeista. Esimerkkejä karbonyyliryhmän sisältävistä yhdisteistä ovat: aldehydi, ketoni, karboksyylihappo, esteri ja amidi. Karbonyyliryhmä antaa näille yhdisteille niille ominaisen kemiallisen luonteen.
Fruktolyysi ja glykolyysi ovat itsenäisiä metabolisia reaktioketjuja (metabolic pathway)
”Suuri osa syödystä glukoosista kulkeutuu maksan läpi luurankolihaksiin, jossa se metaboloituu glykolyysissa ja edelleen sitruunahappokierrossa ensin pyruvaateiksi ja edelleen hiilidioksidiksi (CO2), hapeksi (H2O) ja ATP-molekyyleiksi, tai rasvasoluihin, jossa glukoosimolekyyleistä metaboloidaan glyserolifosfaattia triglyseridien synteesiin ja energiantuotantoon.”
Fruktoosin aineenvaihdunta syntetisoi maksassa glykogeenejä ja de novo lipogeneesissä rasvahappoja ja triglyseridejä.
Tämä synteesi voidaan jakaa kahteen päävaiheeseen:
Trioosien, dihydroksiasetonin (DHAP) ja glyseraldehydin synteesi.
Toisessa vaiheessa trioosit jatkavat aineenvaihduntaa joko glukoneogeneesissä, täyttävät maksan glykogeenejä ja/tai fruktolyysin reaktioketjussa pyruvaatiksi; pyruvaatti muutetaan sitruunahappokierrossa sitraatiksi ja lopuksi de novo synteesissä vapaista rasvahapoista syntetisoidaan palmitiinihappoa.
Palmitiinihappo
Palmitiinihappo on yleisin tyydyttynyt rasvahappo sekä eräs yleisimmistä rakenneosasista eläin- ja kasvirasvoissa. Esimerkiksi ihrassa ja voissa on 25 % palmitiinihappoa. Myös ihmisen rasvasta 25 % on palmitiinihappoa. Palmitiinihapon estereitä ja suoloja kutsutaan palmitaateiksi.
Trioosit ovat kolmesta hiiliatomista muodostuvia monosakkarideja. Dihydroksiasetoni on kaksi hydroksyyliryhmää sisältävä ketoni. Glyseraldehydi on yksinkertaisin monosakkarideista. Se on makea yhdiste, jota syntyy hiilihydraattien hajoamisen seurauksena.
Fruktoosi metaboloituu DHAP:ksi ja glyseraldehydiksi
Fruktoosin aineenvaihdunnan ensimmäinen askel on fruktoosin fosforylaatio, jossa fruktokinaasi muuttaa fruktoosimolekyylin fruktoosi-1-fosfaatiksi. Tämä reaktioketju sitoo fruktoosin aineenvaihdunnan maksaan.
Maksassa esiintyy myös heksokinaasi IV-entsyymiä (Glukokinaasi), joka voi fosforyloida vähäisestä määrästä fruktoosia fruktoosi 6-fosfaattia (glukoneogneettisen reaktioketjun välivaihe). Käytännössä kaikki fruktoosimolekyylit fosforyloidaan maksassa kuitenkin fruktoosi-1-fosfaatiksi.
Kuvakaappaus: Wikipedia
Toisaalta suurin osa glukoosimolekyyleistä jää fosforyloimatta ja kulkee maksan läpi rasvakudokseen ja luurankolihaksiin insuliiniriippuvaisen glukoosinkuljetusmolekyylin (GLUT4) kuljettamana.
Fruktoosi-1-fosfaatti hydrolysoidaan fruktoosi-1fosfaatti aldolaasin (aldolaasi B) avulla dihydoksiasetonifosfaatiksi (DHAP). Dihydroksiasetonifosfaatti on orgaaninen molekyyli, joka esiintyy välituotteena monissa biokemiallisissa reaktioissa, esimerkiksi glykolyysissä. Aldolaasi eli fruktoosi-1,6-bisfosfaattialdolaasi on entsyymi, joka osallistuu glykolyysiin ja glukoneogeneesiin katalysoimalla fruktoosi-1,6-bisfosfaatin reversiibeliä hajoamista glyseraldehydi-3-fosfaatiksi tai glyseraldehydiksi ja dihydroksiasetonifosfaatiksi.
DHAP voi isomerisoitua glyseraldehydi-3-fosfaatiksi, tai glyseroli-3-fosfaatiksi. Glyseraldehydi kinaasi voi muuttaa glyseraldehydin glyseraldehydi-3-fosfaatiksi tai glyseroli-3-fosfaatiksi.
Fruktoosin aineenvaihdunnan tuottamat väliaineet voivat osallistua glukoneogeneesiin ja glykogeenin synteesiin, tai ne voidaan hapettaa pyruvaatiksi ja edelleen laktaatiksi, tai dekarboksyloida asetyylikoentsyymi-A:ksi mitokondrioissa sekä edelleen siirtää vapaiden rasvahappojen synteesiin ja lopulta triglyseridien synteesiin.
Kuva: Fruktoosin aineenvaihdunta dihydroksifosfaatiksi (DHAP), glyseraldehydiksi ja glyseraldehydi-3-fosfaatiksi maksassa.
Glykogeenin synteesi dihydroksiasetonifosfaatista ja glyseraldehydi-3-fosfaatista
Fruktoosin aineenvaihdunta jatkuu glukoneogeneesin lähtöaineista. Fruktoosista metaboloidaan aluksi dihydroksiasetonifosfaattia (DHAP) ja glyseraldehydia fruktokinaasin ja aldolaasi B:n katalysoimana.
Lisääntynyt DHAP- ja glyseraldehydi-3-fosfaatin konsentraatio maksassa ohjaa glukoneogeneesin reaktioketjun kohti glukoosi-6-fosfaattia, glukoosi-1-fosfaattia ja glykogeenin synteesiä.
Fruktoosi on parempi substraatti glykogeenin synteesille kuin glukoosi. Glykogeenivaraston täydennys on etusijalla triglyseridien synteesiin nähden. Kun maksan glykogeenivarasto on täydennetty, fruktoosin ylimääräiset aineenvaihduntatuotteet jatkavat triglyseridien synteesiin.
Triglyseridien synteesi dihydroksiasetonifosfaatista ja glyseraldehydi-3-fosfaatista
Ravinnosta saatu ylimääräinen fruktoosi voidaan muuttaa pyruvaatiksi ja siirtää sitruunahappokiertoon, jossa se muutetaan edelleen sitraatiksi, tai ohjata reaktiosarjaa vapaiden rasvahappojen synteesiin soluliman nestemäisessä sytosolissa. Fruktolyysin syntetisoima DHAP voidaan muuttaa glyseroliksi ja edelleen glyseroli-3-fosfaatiksi triglyseridien synteesiin de novo lipogeneesissa.
”Thus, fructose can provide trioses for both the glycerol 3-phosphate backbone, as well as the free fatty acids in TG synthesis. Indeed, fructose may provide the bulk of the carbohydrate directed toward de novo TG synthesis in humans.”
Runsaasti teollista fruktoosia sisältävän ravinnon on havaittu altistavan hypertriglyseridemialle eli veren liialliselle triglyseridipitoisuudelle.
”The hypertriglyceridemic effects observed are a hallmark of increased dietary carbohydrate, and fructose appears to be dependent on a number of factors including the amount of dietary fructose consumed and degree of insulin resistance.”
Fruktoosin aineenvaihdunnan virheet
Fruktoosin aineenvaihduntaan vaikuttavien kahden tärkeän entsyymin puutos aiheuttaa kaksi synnynnäistä hiilihydraattien aineenvaihdunnan virhettä.
Fruktokinaasin puutos aiheuttaa essentiaalisen fruktosurian. Fruktokinaasi osallistuu reaktioketjuun, jossa fruktoosi muutetaan fruktoosi-1-fosfaatiksi. Tämän entsyymin puutoksen seurauksena fruktoosin aineenvaihdunta jää epätäydelliseksi, jolloin fruktoosia erittyy virtsaan. Fruktosuria on perinnöllinen tila, joka ei kuitenkaan aiheuta kliinisiä oireita, sillä fruktoosi voidaan metaboloida fruktoosi-6-fosfaatiksi heksokinaasin avulla esimerkiksi rasva- ja lihaskudoksissa.
Fruktoosin imeytymishäiriö ja perinnöllinen fruktoosi-intoleranssi
Fruktoosin imeytymishäiriö on ruoansulatuskanavan häiriö, jossa fruktoosin imeytyminen ohutsuolesta verenkiertoon on heikentynyt fruktoosinkuljetusmolekyylien vähäisyyden vuoksi. Imeytymishäiriön oireita ovat mm. vatsakipu, turvotus, ilmavaivat ja ripuli.
Fruktoosin imeytymishäiriö muistuttaa oireiltaan ärtyvän suolen oireyhtymää sekä laktoosi-intoleranssia.
Fruktoosin imeytymishäiriötä ei pidä sekoittaa mahdollisesti hengenvaaralliseen perinnölliseen fruktoosi-intoleranssiin, jossa maksassa fruktoosia pilkkovat entsyymit eivät toimi oikein.
Fruktoosi imeytyy ohutsuolesta ilman ruoansulatusentsyymien apua. Terveen henkilön ohutsuoli pystyy kerrallaan imeyttämään ohutsuolesta verenkiertoon 25-50 grammaa fruktoosia. Fruktoosin imeytymishäiriötä sairastavilla jo alle 25 g fruktoosiannos voi aiheuttaa vatsavaivoja. Sorbitoli voi edelleen heikentää fruktoosin imeytymistä ja lisätä vatsavaivoja. Imeytymätön fruktoosi fermentoituu suolistobakteerien vaikutuksesta ja lisää suolistokaasujen muodostumista.
Monet runsaasti fruktoosia sisältävät hedelmät, esimerkiksi omenat, päärynät, mangot ja vesimelonit, voivat aiheuttaa oireita fruktoosin imeytymishäiriötä sairastavalla. Oireita voi tulla myös runsaasti fruktoosia sisältävästä pöytäsokerista, hunajasta, maissisiirapista, rusinoista, hedelmämehuista ja fruktaaneista (FODMAP).
Fruktoosin imeytyshäiriöön ei ole parantavaa hoitoa, mutta sen aiheuttamia oireita voi välttää minimoimalla fruktoosin saannin. Myös FODMAP-ruokavalio voi auttaa oireiden helpottamisessa. Lähde: Wikipedia
Perinnöllinen fruktoosi-intoleranssi (HFI)
Perinnöllinen fruktoosi-intoleranssi (HFI) johtuu synnynnäisestä aldolaasi B-entsyymin puutoksesta. Puutoksen seurauksena sakkaroosi, fruktoosi ja sorbitoli aiheuttavat oireita synnynnäistä fruktoosi-intoleranssia sairastavilla. Aldolaasi-B entsyymin puutos johtaa fruktoosi-1-fosfaatin kerääntymiseen maksasoluihin, munuaisiin ja ohutsuoleen. Ajan mittaan tämä johtaa maksasolujen tuhoutumiseen. HFI vaikuttaa myös glukoneogeneesiin, glykogenolyysiin ja adenosiinitrifosfaatin (ATP) regeneraatioon. HFI aiheuttaa mm. pahoinvointia, oksentelua, kouristeluja, ärtyisyyttä, hypoglykemiaa, keltatautia, verenvuotoa, maksan liikakasvua sekä mahdollisesti munuaisten vajaatoimintaa. HFI voi johtaa kuolemaan, mutta se on melko harvinaista.