Kuinka ravinto ja elintavat vaikuttavat MS-taudin etenemiseen? Osa 3:
Kuinka ravinto ja elintavat vaikuttavat MS-taudin etenemiseen? Osa 3 jatkaa ravinnon ja elintapojen vaikutuksen selvittämistä MS-taudin ja siihen assosioituvien oireiden taustalla.
Mitä eroa on infektiolla ja inflammaatiolla?
Autoimmuunitaudin voi laukaista jokin infektio, kuten Epstein-Barr- tai herpesvirus. Inflammaatio altistaa sairastumiselle ja pahentaa immuunivälitteisten tulehduksellisten sairauksien oireita. Eräät ravintoaineet aiheuttavat oksidatiivista stressiä, joka ylläpitää ja pahentaa inflammaatiota.
Infektio ja inflammaatio menevät helposti sekaisin, koska molemmat kertovat tulehduksesta. Ne eivät kuitenkaan tarkoita samaa asiaa.
Infektion aiheuttama lyhytaikainen tulehdus (tartuntatulehdus) auttaa elimistöön tunkeutuneen sairastuttavan mikrobin tuhoamisessa. Jatkuva matala-asteinen tulehdus (inflammaatio) on kudoksia ärsyttävä tila, joka voi kehittyä mm. vamman, ravinnon (postbrandiaalinen tulehdus), tupakoinnin, alkoholin ja eräiden toksiinien sekä joidenkin tuntemattomien syiden seurauksena, kuten eräät autoimmuunitulehdukset (esimerkiksi reuma).
Matala-asteinen tulehdus ei tavallisesti näy ulospäin tai oireile kipuna. Tutkimukset viittaavat siihen, että matala-asteinen tulehdus on kuitenkin lähes kaikkien kroonisten tautien osatekijä.
”Ne ruoka-aineet, jotka vähentävät tulehdusta tuntuvat edistävän ihmisen terveyttä muutoinkin. Ruokavalio, joka vähentää voimallisesti tulehdusta, vähentää myös kolesterolia, verenpainetta, aterian jälkeistä hapetusstressiä, LDL-kolesterolin hapettumista ja verensokeria aterian jälkeen ja paastossa sekä estää lihomista.” – Pronutritionist
Infektio siis puolustaa elimistöä mikrobeja vastaan. Pitkään jatkuva matala-asteinen inflammaatio on elimistölle haitallinen tila, koska se voi aiheuttaa kudosvaurioita. Wikipedian mukaan autoimmuunitulehdus vahingoittaa elimistöä immuunipuolustuksen hyökätessä elimistön omia soluja vastaan.
C-reaktiivinen proteiini eli CRP
Elimistön tulehduksista kertoo verinäytteestä mitattava CRP eli C-reaktiivinen proteiini. CRP on maksan syntetisoima akuutin infektion proteiini, joka sitoutuu solun erilaisiin ainesosiin, kuten polysakkarideihin, lipideihin, nukleiinihappoihin, nukleotideihin sekä kationeihin kuten hepariiniin, protamiiniin ja histoineihin.
CRP on komplementtijärjestelmän aktivoija, joka edistää vierasaineiden opsonisaatiota ja fagosytoosia. CRP osallistuu luontaiseen immuniteettiin ja vierasaineiden eliminointiin. Opsonisaatio on prosessi, jossa infektoivan patogeenin pintaan tarttuu siihen erikoistunut vasta-ainemolekyyli eli opsoniini, jonka avulla syöjäsolut (fagosyytit) tunnistavat ja tuhoavat patogeenit.
CRP:n pitoisuus veressä nousee bakteeri-infektioiden ja muiden tulehdustilojen sekä kudosvaurion yhteydessä nopeasti. CRP:n normaali viitealue on alle 10 mg/l, mutta infektion aikana CRP:n määrä voi kasvaa jopa 1000-kertaiseksi viitealueeseen verrattuna.

Ruokavalio, laihtuminen ja CRP
Lihavuus on matala-asteisen inflammaation yksi tärkeä syy. Laihtuminen voi laskea inflammaatiota mittaavaa CRP-arvoa jopa 80 %. Myös ruokavalio vaikuttaa tulehdusta mittaavaan CRP-arvoon. Terveellinen ruoka, kuten kasvikset, marjat, hedelmät ja kala voivat laskea tulehdusarvoja jopa kolmanneksella.
Wikipedia kertoo, että jo muutaman päivän vesipaasto vahvistaa kehon immuunijärjestelmää taistelussa tulehduksia vastaan. Vastaavia tuloksia on saatu kerran tai kahdesti kuussa toteutettavilla nelipäiväisillä niukan ravinnon jaksoilla.
Elimistö näyttäisi pääsevän paaston ja niukan dieetin avulla eroon immuunijärjestelmän vahingoittuneista ja vanhentuneista osista, joka johtaa immuunijärjestelmän uusiutumiseen (Kari Tyllilä: Yllättävä löytö voi tuoda apua syöpähoitoihin: Paasto uudistaa immuunijärjestelmää).
Tulehdus ja sytokiinit
Sytokiinit ovat immuunijärjestelmän säätelijöitä. Ne ohjaavat immuunijärjestelmän kaikkien solujen erilaistumista, kasvua ja toiminnallista säätelyä. Sytokiini on yleisnimitys yli sadalle pienimolekyyliselle proteiinirakenteiselle välittäjäaineelle.
Valkosolut tuottavat sytokiineihin lukeutuvia interferoneja virusinfektion aikana. Interferonien tarkoituksena on estää virusten lisääntyminen infektion alkuvaiheessa. Proinflammatoristen eli inflammaatiota lisäävien interferonien (sekä happiradikaalien) ensisijaisena tehtävänä on tappaa elimistöön päässeitä viruksia, bakteereita ja ja sieniä.
Jos immuunivasteeseen osallistuvia sytokiineja tai happiradikaaleja muodostuu elimistössä liikaa, ne vaurioittavat kudoksia ja altistavat sairastumiselle.
Beetainterferoneja käytetään MS-taudin oireita hillitsevänä lääkkeenä. Toisaalta gammainterferoni, jota naisilla muodostuu luonnostaan miehiä enemmän, assosioituu suurina pitoisuuksina MS-taudin puhkeamiseen ja pahenemiseen.
Tärkeät sytokiinit
Kuva sytokiinien merkityksestä ja hierarkkisista säätelyverkoista tarkentuu koko ajan. Sytokiineja on tunnistettu yli sata. Pelkästään interleukiiniperheeseen kuuluvia sytokiineja tunnetaan 29. Sytokiinien tutkimus tarjoaa uusia lähestymistapoja myös autoimmuunitautien ja syöpien hoitoon.
Immuunijärjestelmän kannalta keskeisiä sytokiinejä ovat interleukiinit (IL), interferonit (IFN), tuumorinekroositekijä alfa (TNF-α), ja solutyyppispesifiset kasvutekijät, kuten granulosyyttikasvutekijä (G-CSF) ja erytropoietiini (EPO).
Sytokiinien eritys lisääntyy infektion aikana, mikä vahvistaa elimistön puolustautumista taudinaiheuttajia vastaan. Immuunivasteeseen ja infektion torjuntaan osallistuvat ainakin proinflammatoriset sytokiinit, kuten IL-1, IL-6, TNF- α.
IL-17 on nopeasti kasvava sytokiiniperhe, jonka jäsenet eroavat rakenteellisesti muista sytokiineista. IL-17 on keskeinen sytokiini MS-taudin patogenesissä (Gold & Lühder, Interleukin-17 – Extended Features of a Key Player in Multiple Sclerosis).
Monet sytokiinit aiheuttavat ja ylläpitävät elimistön matala-asteista tulehdusta. Toisaalta sytokiinit voivat olla myös inflammaatiota vähentäviä eli anti-inflammatorisia, kuten mm. IL-4, IL-10 ja TGF- β.
Sytokiinien säätelyverkot
Sytokiinit muodostavat toiminnalllisia verkostoja. Yksittäinen sytokiini vaikuttaa tavallisesti useisiin solutyyppeihin, mutta sen aikaansaamat vasteet eri soluissa voivat olla täysin erilaiset.
Monissa immuunivälitteisissä tulehduksellisissa sairauksissa aktivoituu osin samanlainen sytokiiniverkosto, mutta yksittäisen sytokiinin merkitys eri sairauksien patogeneesissa voi vaihdella paljonkin. Yhtenä esimerkkinä tuumorinekroositekijä (TNF-α), joka vaikuttaa mm. nivelreuman, selkärankareuman, tulehduksellisten suolistotautien ja psoriaasin patogeneesissa.
Sytokiinit toimivat ajallisesti ja paikallisesti tarkan säätelyn alaisina hierarkkisina säätelyverkostoina. Jos säätely jostakin syystä pettää, sytokiinien ylituotanto voi toimia laukaisevana mekanismina monissa sairauksissa, kuten autoimmuunitaudeissa.
Sytokiinireseptoreiden signalointi
Sytokiinien biologiset vaikutukset välittyvät solun pinnalla sijaitsevien erityisten reseptoreiden kautta. Interferonien, useimpien interleukiinien ja solutyyppispesifisten kasvutekijöiden (EPO, TPO, GM-CSF, G-CSF) reseptorit välittävät vaikutuksensa hematopoieettiseen sytokiinireseptoriperheeseen kuuluvien reseptoreiden kautta.
Kaikkien reseptroreiden aktivaatiomekanismi on samankaltainen: sytokiinin sitoutuminen reseptorin solunulkoiseen osaan saa aikaan ketjuen pariutumisen ja johtaa reseptoriin kiinnittyneiden JAK-tyrosiinikinaasien (JAK1-3 ja TYK2) aktivaatioon ja signaalinvälitykseen erikoistuneiden proteiinien fosforylaatioon sekä muutoksiin mm. DNA-synteesissä ja transkriptiossa (Levy ja Darnell 2002, O’Shea ym. 2002).
Luettavaa sytokiineista
- Researchers Identify 2 Cytokines Responsible for Chronic Flares in Autoimmune Diseases
- Cytokines in Multiple Sclerosis – Possible Targets for Immune Therapies
- Cytokine Profile in Patients with Multiple Sclerosis Following Vitamin D Supplementation
- Uutta sytokiineistä, Olli Silvennoinen ja Mikko Hurme
- Wikipedia: Cytokine
- Cytokines, Inflammation and Pain
Kuinka elimistö reagoi tulehdukseen?
Elimistö reagoi tulehdukseen tavallisesti verisuonimuutoksilla sekä kudosnesteen ja tulehdussolujen kertymisellä tulehdusalueelle. Verisuonimuutosten seurauksena hiussuonten seinämät muuttuvat läpäisevimmiksi ja tulehdusalueelle kertyy proteiineja ja nestettä.
Tulehdusreaktio houkuttelee paikalle myös valkosoluja, kuten syöjäsoluja, joiden tehtävänä on puolustaa elimistöä ulkoisilta taudinaiheuttajilta ja siivota tulehdusaluetta vaurioituneista soluista. Tulehduksien aiheuttama ”märkä” muodostuu tulehdussoluista, taudinaiheuttajista, osin tuhoutuneesta kudoksesta ja kudosnesteestä.
Tulehduksen oireet ovat rubor, tumor, calor, dolor ja functio laesa eli punoitus, turvotus, kuumotus, kipu ja toimintakyvyn heikkeneminen.
Ravinto ja inflammaatio
Pitkään jatkuva matala-asteinen tulehdus kasvattaa sairastumisen riskiä. Tutkimusten mukaan inflammaatio on useimpien kroonisten sairauksien taustatekijä. Inflammaatio altistaa mm. autoimmuunitaudeille, sydän- ja verisuonitaudeille, syöville, tyypin 2 diabetekselle, lihavuudelle ja Alzheimerin taudille.
Rasvakudos erittää runsaasti erilaisia tulehdussytokiineja, joten lihavuus ylläpitää ja lisää inflammaatiota. Laihduttaminen voi merkittävästi vähentää elimistöä rasittavaa matala-asteista tulehdusta.
Aterianjälkeinen (postbrandiaalinen) verensokerin nousu kasvattaa oksidatiivista stressiä muodostamalla happiradikaaleja. Oksidatiivinen stressi pahentaa inflammaatiota. Mitä korkeammaksi verensokeri nousee, sitä enemmän muodostuu happiradikaaleja.
Ravinto vaikuttaa inflammaatioon monin tavoin.
Ravinto voidaan jakaa karkeasti tulehduksia aiheuttaviin, neutraaleihin ja tulehduksia hillitseviin ravintoaineisiin. Ravintoaineiden aiheuttamaan tulehdusvasteeseen vaikuttavat mm. ravinnon määrä ja muut samaan aikaan nautitut ravintoaineet.
Rasvat ovat tavallisilla annoksilla tulehduksen kannalta yleensä neutraaleja. Värikkäiden marjojen, hedelmien ja kasvisten syöminen lievittää tulehdusta. Imeytymättömät proteiinit voivat lisätä suoliston tulehduksia, mutta lihan, kanan ja äyriäisten tulehdusvaikutuksista on hyvin vähän tutkittua tietoa.
Tulehduksia vähentäviä (anti-inflammatorisia) ruokia ovat mm.
- Rasvainen kala
- Neitsytoliiviöljy
- Kala
- Mantelit ja pähkinät
- Marjat ja hedelmät
- Monet kasvikset
- Appelsiinimehu
- Granaattiomena
- Kaakao
- Punaviini
Inflammaation kannalta neutraaleja ruokia ovat mm.
- Rypsiöljy
- Margariini
- Voi
- Soija ja palkokasvit
- Meijerituotteet
- Kananmunat
- Eräät täysjyvätuotteet
- Monet kasvikset
- Tumma pasta
- Peruna
- Leipä
- Vihreä tee
- Kahvi
- Valkoviini
- Maito ja piimä
Tulehduksia lisääviä ruokia ovat mm.
- Kerma suurina annoksina
- Makkarat ja lihajalosteet
- Runsas sokeri
- Runsas fruktoosi (fruktoosisiirappi?)
Lue lisää:
Pronutritionist: Anti-inflammatorinen eli tulehdusta vähentävä ruokavalio

Inflammaation vaikutus RRMS- ja PPMS-tautien oireisiin
Tulehdustekijät ovat havaittavissa aaltoilevasti etenevän RRMS-taudin patologiassa ja assosioituvat selkeästi taudin oireisiin. RRMS-tautimuodossa keskushermostossa ilmenevät tulehduspesäkkeet (leesiot) ovat yhteydessä taudinkuvaan liittyviin kliinisiin pahenemisvaiheisiin. Inflammaation helpottuminen ilmenee remissiona, jolloin taudin oireet paranevat joko osittain tai lähes täysin etenkin taudin varhaisvaiheessa.
Progressiivinen MS-tauti
Ensisijaisesti etenevä MS-tauti (primaarisprogressiivinen, PPMS) on MS-taudin alatyyppi, jossa oireet ja invaliditeetti lisääntyy sairauden alusta alkaen tasaisesti ilman selviä inflammaatioon assosioituvia pahenemisvaiheita, RRMS etenee toissijaisesti eteneväksi (SPMS) taudiksi yleensä noin parin vuosikymmenen aikana. SPMS ja PPMS muistuttavat hyvin läheisesti toisiaan.
”PPMS-potilailla yleisimmät taudin alkuoireet olivat motoriset-, pikkuaivo- ja tuntohäiriöt. Motorisen toiminnan häiriöt olivat yleisimmät löydökset kliinisessä neurologisessa tutkimuksessa. Kaikilla PPMS-potilailla oli virtsaustoiminnan häiriöitä, joista tihentynyt virtsaamistarve ja siihen liittyvä virtsan karkailu olivat yleisimmät oireet. Urodynaamisen tutkimuksen yleisimmät löydökset olivat virtsarakon seinämälihaksen yliaktiivisuus (detrusor hyperrefleksia) sekä seinämälihaksen ja virtsaputken sulkijalihaksen koordinoimaton supistelu (detrusor sphinkterin dyssynergia, DSD).” – Maritta Ukkonen: Primaarisprogressiivinen MS-tauti, kliininen, immunologinen ja radiologinen kuva
Tulehduksen vaikutusta ei ole poissuljettu myöskään etenevässä MS-taudissa. Inflamaation voi aiheuttaa autoimmuunitulehdus tai solujen (oligodendrosyyttien) rappeutumisen eli sytodegeneraation aiheuttama neurologinen tulehdus.
”Adheesiomolekyylien ja joidenkin sytokiinien ilmentymisen lisääntyminen viittaa siihen, että tulehduksellista aktiviteettia esiintyy pidemmälle edenneessä PPMS-taudissakin.” – Maritta Ukkonen: Primaarisprogressiivinen MS-tauti, kliininen, immunologinen ja radiologinen kuva
Tutkimukselliset löydöt
Etenevissä MS-taudeissa on havaittavissa runsaasti molekyyli- ja solutason muutoksia, jotka selittävät taudinkuvaan liittyvää neurologista rappeutumista (neurodegeneraatiota). Tällaisia neurologiseen rappeutumiseen assosioituvia muutoksia ovat mm.
- keskushermoston syöjäsoluina toimivien mikrogliasolujen aktivoituminen
- kroonisen hapettumisreaktion aiheuttamat vauriot keskushermoston soluissa
- mitokondrioihin kumuloituvat vauriot keskushermoston viejähaarakkeissa
- ikään liittyvä atrofia
- viejähaarakkeiden signaalinvälityksen havaittava heikkeneminen.
Tällaiset patologiset muutokset voivat johtua autoimmuunitulehduksen aiheuttamista viejähaarakkeiden eristekalvojen vaurioista (demyelinaatio), mutta syynä voi olla myös tautiin liittyvä keskushermoston solujen (neuronien ja oligodendrosyyttien) primaari rappeutuminen.
Mahdollisesti moolemmat, sekä inflammaatio että keskushermoston solujen rappeutuminen (sytodegeneraatio) vaikuttavat etenevien MS-tautimuotojen patogeneesiin.
Patologiset mekanismit, jotka ylläpitävät neurodegeneraatiota ja aiheuttavat PPMS-ja SPMS-potilaille kudosvaurioita, tunnetaan huonosti. Nämä tekijät liittyvät ilmeisesti perifeerisen immunologisen toleranssin virheelliseen toimintaan.
Taudin aiheuttamasta neurodegeneraatiosta on esitetty (ainakin) kaksi hypoteessia: inside-out-hypoteesi ja outside-in-hypoteesi.
Inside-out hypoteesin mukaan taudin alusta alkaen etenevä keskushermoston solujen rappeutuminen on kaikkien tautiin liittyvien prosessien selittävä tekijä.
Outside-in hypoteesi olettaa, että taudin varhaisvaiheessa ilmenevät inflammaatioon assosioituvat demyelinoivat prosessit laukaisevat joukon keskushermostoa rappeuttavia tapahtumaketjuja.

Osallistuuko suoliston mikrobiomi autoimmuunitaudin patogeneesiin?
Viime aikoina on saatu viitteitä siitä, että suoliston mikrobiomin hyvinvoinnilla on tärkeämpi rooli etenevän MS-taudin taudinkuvassa kuin on aiemmin oletettu.
Tieto mikrobiomista ja sen merkityksestä isäntäorganismille täsmentyy koko ajan. Vagus-hermo välittää tietoa ruoansulatuselimistön tapahtumista aivoille. Se toimii suorana välittäjänä mikrobiomin ja keskushermoston välillä.
Mikrobiomi vaikuttaa keskushermostoon muokkaamalla signaalireittejä aivo-suolisto-akselilla. Tämä kaksisuuntainen kommunikaatioverkko hermoston ja suoliston välillä aktivoi hermoston makrofageja ja vaikuttaa neurologisiin tapahtumiin säätelemällä hermoston immuuniaktiivisuutta.
Mikrobiomin merkitys
BBC kirjoittaa, että kehon solujen kokonaismäärästä 43 % on ihmisen omia soluja. Suurin osa kehossamme elävistä soluista kuuluu kuitenkin mikrobiomin bakteereille, arkeille, viruksille ja sienille.
Ihmisen DNA:ssa on noin 23 000 proteiineja koodaavaa geeniä. Geenit säätelevät solujemme, kudostemme ja elimistömme rakennetta. Geenien väliset alueet ohjaavat geenien toimintaa. Oman genomin lisäksi kehossamme on mikrobiomin geneettistä materiaalia, joka koostuu 2-20 miljoonasta geenistä.
DNA, mutaatiot ja yhden emäksen variaatiot
Ihmisen DNA on noin 3 miljardia emästä pitkä kaksoisjuoste. DNA:n rakenteessa toistuu neljä emästä, joita kuvataan kirjaimilla A (adeniini), T (tymiini, C (sytosiini) ja G (guaniini). A ja T sekä C ja G muodostavat DNA:n kaksoisjuosteessa emäspareja.
Geenit eli perintötekijät muodostuvat eri mittaisista DNA-jaksoista
Solun jakautuminen edellyttää DNA:n kahdentumista. Prosessi on hyvin täsmällinen, mutta aika ajoin siinä tapahtuu virheitä ja DNA-juosteen alkuperäinen emäsjärjestys muuttuu. Tällaiset virheet aiheuttavat geenimutaatioita.
Geenimuutosten kolme lähdettä ovat vanhemmilta saatu perimä, elintapojen ja ympäristön tuoma altistus (myrkyt, patogeenit, ravinto jne.) sekä sattumanvaraiset DNA:n kopioitumisvirheet. Kopioitumisvirheitä tapahtuu jatkuvasti. Aina, kun solu jakaantuu, aiheutuu DNA:han keskimäärin kolme virhettä. Tällaiset geenimutaatiot voivat käynnistää syövän.
Pistemutaatiot eli yhden emäksen variaatiot (Single Nucleotide Polymorphism), joissa esimerkiksi DNA:n emäsjuosteen jonkin geenin emäsparissa adeniini muuttuu sytosiiniksi, ovat hyvin yleisiä. Yleensä nämä ovat neutraaleja, mutta jotkin yhden nukleotidin polymorfismit assosioituvat lisääntyneeseen sairastumisriskiin.
Toinen genomi
Professori Sarkis Mazmanian (Caltech) kertoi BBC:lle, että periaatteessa meillä on kaksi toisiinsa vuorovaikuttavaa genomia. Ne ”kommunikoivat” keskenään kemiallisten signaalien välityksellä. Tällaisia mikrobiomin tuottamia hermostoon vaikuttavia välittäjäaineita ova esimerkiksi eräät mikrobien aineenvaihduntatuotteet, kuten dopamiini, serotoniini ja GABA.
Mikrobiomin tuottamat kemialliset signaalit voivat vaikuttaa myös epigeneettisesti ihmisen omaan genomiin. Tämä tapahtuu esimerkiksi siten, että johonkin geenin emäksistä kiinnittyy ympäristötekijöiden säätelemänä geenin transkriptioon vaikuttava metyyliryhmä.
Mikrobiomiin vaikuttavat ympäristötekijät voivat olla viruksia, bakteereita, sieniä, tietyn kemiallisen koostumuksen omaavia ravintoaineita sekä toksisia tai inflammatorisia kemikaaleja. Nämä voivat heikentää immuunijärjestelmän säätelyä ja edesauttaa epigeneettisten muutosten, pistemutaatioiden ja geenimutaatioiden kehittymistä DNA:han.
Yhden nukleotidin polymorfismit (single nucletide polymorphism, SNP) assosioituvat moniin sairauksiin, kuten syöpiinn ja autoimmuunitauteihin. Esimerkiksi tyypin 1 diabeteksessa ja MS-taudissa tällaisia tautiin assosioituvia yhden nukleotidin polymorfismeja on tunnistettu muun muassa geenin CYP27B1 eri lokuksissa.
Geenit eivät ole täysin muuttumattomia. Ympäristötekijät vaikuttavat geenien toimintaan.
Myös epigeneettinen muutos, jossa geenin yhden tai useamman emäksen päälle on kiinnittynyt metyyliryhmä vaikuttaa geenin ekspressioon ja transkriptioon.

Kuinka mikrobiomi vaikuttaa elimistöön?
Suoliston mikro-organismit estävät vieraiden ja mahdollisesti haitallisten mikrobien pesiytymisen suolistoon ja pääsyn suoliston kautta verenkiertoon.
Mikrobiomi vaikuttaa myös ruoansulatukseen, aineenvaihduntaan, immuunijärjestelmän säätelyyn sekä eräiden vitamiinien ja muiden tärkeiden yhdisteiden, kuten dobamiinin, GABAn ja serotoniinin synteesiin ja edelleen keskushermoston toimintaan mm. vagus-hermon välityksellä.
Onko antibiooteilla ja rokotuksilla vaikutuksia mikrobiomiin?
Antibiootit ja rokotukset ovat pelastaneet kymmeniä tai satoja miljoonia ihmishenkiä viimeisen vuosisadan aikana, mutta joidenkin tutkijoiden mukaan mikrobiomin lajikirjo on pienentynyt infektioilta suojaavan taistelun seurauksena ja tämä on heikentänyt mikrobiomin vaikutusta immuunijärjestelmän säätelyyn.
Hypoteesin mukaan mikrobiomin lajikirjon pienentyminen vaikuttaa immuunijärjestelmän säätelyn kautta sairastumissalttiuden lisääntymiseen. Erityisesti sairastumisalttiuden lisääntyminen vaikuttaa allergioihin ja autoimmuunitauteihin.
Professori Ruth Ley (Max Planck Institute) totesi BBC:lle, että vaikka olemmme taistelleet menestyksekkäästi infektioita vastaan, autoimmuunitautien ja allergioiden määrä on kääntynyt selvään kasvuun.
Tulkitsen tämän niin, että koska mikrobiomi yleensä periytyy äidiltä lapselle, voivat pienet mikrobiomin lajikirjon muutokset kumuloitua sukupolvien aikana ja heikentää pitkällä aikajänteellä immuunijärjestelmän säätelyä. Se voisi selittää väestötasolla eräiden tautien yleistymisen.
Rokotukset ja antibiootit eivät kausaalisesti aiheuta autoimmuunitauteja, mutta ovat voineet useiden sukupolvien aikana vaikuttaa autoimmuunitautien kehittymisen kannalta otollisemman immunologisen ympäristön rakentumiseen. Tällainen spekulaatio kuulostaa ihan järkeenkäyvältä.
Näkökulma: Rokotteiden sisältämät viruksen proteiinit toimivat autoimmuunitaudin laukaisijoina minimaalisen pienellä todennäköisyydellä, mutta näin kävi surullisessa narkolepsiaepidemiassa. Yleisesti ottaen rokotteet ovat hyvin turvallisia. Virus, jolta rokote suojaa voi laukaista autoimuunitaudin myös rokottamattomilla.
Rokottaminen voi laukaista vakavan allergisen reaktion tai sairauden, mutta todennäköisyys sellaiselle on häviävän pieni. Myös rokotteen sisältämien tehoste- ja säilöntäaineiden pelko on aiheeton; hengittämällä elimistöön kulkeutuu taajama-alueilla jo yhdessä päivässä rokotteisiin verrattuna moninkertainen määrä teollisuudesta ja liikenteestä peräisin olevia haitallisia mikropartikkeleita. Hengitysilman pienhiukkaset kulkeutuvat keuhkoista verenkiertoon ja vaikuttavat siten terveyteen.
Maailmanlaajuisesti ilmansaasteet tappavat vuosittain miljoonia ihmisiä. Suurin ongelma on Aasiassa ja Afrikassa. Tämä on rokotteita todellisempi ja akuutimpi uhkakuva myös Euroopassa.
Teollinen ruoka yksipuolisti mikrobiomia
Mikrobiomin heikentymiseen on vaikuttanut myös viime vuosisadalla alkanut ravinnon teollistuminen. Teollisesti valmistetut vähemmän ravinteita ja enemmän energiaa sisältävät ruoat ja rasvat sekä runsas sokereiden käyttö ovat syrjäyttäneet luonnnollisemmat ravinnonlähteet.
Lihan ja sokereiden määrä ravinnossa on lisääntynyt samaan aikaan, kun hapatettujen ruokien ja kasvisten saanti on vähentynyt. Punainen liha, lihajalosteet, transrasvat ja sokerit assosioituvat tutkimuksissa heikentyneen suolistoterveyden ja suoliston tulehdusten kanssa; nämä heikentävät immuunijärjestelmää ja sen säätelyä.

Punainen liha ja suoliston terveys
Runsaan proteiinien saannin kohdalla ongelmia aiheuttaa se, että vaikka proteiinit pilkotaan tärkeiksi aminohapoiksi ja peptideiksi ohutsuolessa, osa proteiineista ei imeydy ohutsuolesta elimistön hyödynnettäväksi, vaan päätyy paksusuoleen, jossa ne ravitsevat mikrobiomin huonoja bakteereita.
Imeytymättömän proteiinin vaikutuksesta paksusuoleen syntyy imeytymätöntä rautaa, ammoniakkia, amiineja, sulfideja ja haaraketjuisia rasvahappoja (BCFA).
Erityisesti lihan paistamisen yhteydessä Mailard-reaktiossa (ruskistumisessa) syntyy sokeroituneita proteiineja, jotka eivät imeydy ohutsuolessa, vaan kulkeutuvat paksusuolen bakteerien fermentoitavaksi (Tuohy et al. 2006). Lähde: Pronutritionist
Ravitsemuksessa tapahtunut muutos ei tietenkään ole yksiselitteisesti huono asia. Ravintoa on enemmän ja monipuolisemmin tarjolla kuin koskaan aiemmin historiassa. Samaan aikaan pikaruoka- ja herkuttelukulttuurilla on kuitenkin hintansa: immuunijärjestelmän toiminnan säätelyyn osallistuvan mikrobiomin heikentyminen on ehkä mahdollistanut aiemmin harvinaisten tautien ja oireyhtymien yleistymisen.
Autoimmuunitautien, allergioiden ja autismin lisääntyminen voisi siis selittyä väestötasolla tapahtuneilla mikrobiomin pitkän aikavälin muutoksilla. Tämä on mielenkiintoinen ajatus.

Ymäristömuuttujat ja terveys
Evoluutio on tehnyt meistä ympäristön muutoksiin hyvin sopeutuvan lajin. Ympäristön muuttuminen mm. ravinnon ja erilaisten kemikaalien osalta on nykyään kuitenkin niin nopeaa, ettei ihmisen aineenvaihdunta ja immuunijärjestelmä ehdi sopeutua muutoksiin.
Kun ihmiset aiemmin sairastuivat ja kuolivat infektioihin, nyt infektioita suurempia uhkia ainakin kehittyneissä maissa ovat elintapoihin assosioituvat kardiometaboliset oireyhtymät, sydän- ja verisuonitaudit, diabetes, syövät jne.
Ravintoaineiden puutokset ja ympäristön myrkyt altistavat sairastumiselle
Välttämättömien ravintoaineiden puutos ei välittömsti johda sairastumiseen, sillä keho varastoi jonkin verran välttämättömiä vitamiineja ja mineraaleja. Elimistössä on simerkiksi B12-vitamiinia yleensä riittävästi kattamaan muutaman vuoden tarpeen, vaikka sitä ei ravinnosta saisikaan. Vakavien puutosoireiden kehittyminen edellyttää pidempiaikaista vitamiinien tai mineraalien puutosta.
Elimistöllä on myös monia aineenvaihduntamekanismeja elintoimintoja ylläpitävien elinten energiansaannin turvaamiseksi. Solut saavat energiaa hiilihydraateista, rasvoista ja proteiineista.
Kun ravintoa ei ole saatavilla, elimistö muuttaa varastorasvoja ketoaineiksi ja glukoneogeneesissä ketoaineita edelleen glukoosiksi tai soluissa energiaksi. Kun elimistön glykogeenit ja rasvavarastot loppuvat, elimistö alkaa tuottaa ketoaineita vapaista proteiineista ja rasvahapoista. Ravinnon jatkuva puutos saa aineenvaihdunnan pilkkomaan lihaksia aminohapoiksi, joita voi käyttää ketoaineina. Näiden selviytymismekanismien ansiosta terve ihminen voi elää jopa kuukauden pelkällä vedellä.
Toksisten aineiden kumuloituminen elimistöön ja välttämättömien ravinteiden puutokset altistavat kuitenkin pitkään jatkuessaan sairastumiselle.
Ravinto ja suolisto
Yksipuolinen ravinto, liiallinen hygienia, runsas alkoholi, tupakointi sekä eräät lääkkeet voivat heikentää suoliston mikrobiomia. Tälla on vaikutuksia terveyteen, koska suoliston mikribiomia tarvitaan mm. suojaamaan suolistoa ulkoisilta taudinaiheuttajilta, vähentämään suolistotulehdusten vaaraa, ehkäisemään suolistosyöpää ja pilkkomaan ravinnon sulamattomia kuituja.
Monista hedelmistä, kasveista, marjoista, tummasta suklaasta ja kahvista saatavilla polyfenoleilla on suoliston mikrobiomille ja painonhallinnalle ilmeisen myönteisiä vaikutuksia. Ne tukevat suoliston terveyttä ylläpitävien bifidobakteerien kasvua. Punaisesta lihasta saatava hemirauta voi pahentaa suoliston tulehduksia, mutta samaan aikaan saatava resistentti tärkkelys vähentää inflammaatiota.

RRSM ja PPMS
MS-taudin kaksi yleisintä mutoa ovat taudinkuvaltaan ja patologisilta mekanismeiltaan hyvin erilaisia tauteja. On ehkä aiheellista harkita sellaista vaihtoehtoa, että RRMS ja PPMS ovat kaksi erillistä sairautta tai monitekijäistä oireyhtymää.
Ne muistuttavat monin tavoin toisiaan, mutta näiden kahden MS-taudin patogeneesi poikkeaa toisistaan merkittävällä tavalla. RRMS on tulehduksellinen autoimmuunitauti, jossa keskushermoston tulehdukset laukaisevat MS-tudille ominaisen demyelinoivan autoimmuunireaktion. PPMS on sairauden alusta alkaen neurodegeneratiivinen, hermoston soluja rappeuttava sairaus, jossa oligodendrosyyttien tuhoutumista ja atrofiaa tapahtuu tasaisesti ilman inflmaatioon assosioituvia pahenemisvaiheita.
Tällaista hypoteesia tukee kliinisten löydösten ohella myös se, että anti-inflammatoriset ja immunosupressiiviset lääkkeet eivät toimi toivotulla tavalla etenevissä MS-taudeissa, vaikka nillä saadaan hyviä hoitotuloksia aaltoilevaa tautimuotoa sairastavilla.
Myöskään kantasoluhoidosta ei löydy apua etenevään MS-tautiin. Kantasoluhoidossa potilaalta kerätään kantasoluja, joita kasvatetaan petri-maljoissa. Kantasolujen keräämisen jälkeen potilaan virheellisesti toimiva immuunijärjestelmä tuhotaan voimakkaalla kemoterapialla. Viimeisessä vaiheessa kantasoluista istutetaan potilaalle uusi immuunijärjestelmä.
Kaiken kaikkiaan kantasoluhoito kestää noin kuukauden ja sillä on saatu hyviä hoitotuloksia RRMS-potilailla. PPMS- ja SPMS-potilaille kantasoluhoito ei ainakaan nykyisellään sovellu.
Immuunijärjestelmää hillitsevillä lääkkeillä ja kantasoluhoidolla ei ole toivottua vaikutusta etenevässä MS-taudissa, koska immuunijärjestelmän virheellinen toiminta ei ole oireiden ensisijainen syy. Etenevä MS-tauti ei myöskään ole ensisijaisesti tulehduksellinen sairaus, koska tulehduksia vähnetävillä lääkkeillä ei saada toivottua vastetta.
Tästä hypoteesista ei vallitse tietellistä konsensusta, mutta etenevien MS-tautien tutkimus on lisääntynyt ja viime aikoina on saatu selkeitä viitteitä siitä, että PPMS on osin virheellisesti ymmärretty sairaus; sen sekoittaminen relapsoivaan-remittoivaan MS-tautiin vain pahentaa tilannetta ja hidastaa tutkimustyötä.
Niin tai näin, molemmissa MS-taudin muodoissa ravinto ja elintavat vaikuttavat taudin etenemiseen, mutta erilaisten patologisten prosessien ja aineenvaihduntakanavien kautta.
Ehkäpä MS-taudin yksilölliset oireet ja taudinkulku eri potilailla selittyy sillä, että kahteen yleisimpään MS-taudin muotoon vaikuttavat erilaiset geenivariaatiot, geenien alleelit, yhden nukleotidin polymorfismit ja epigeneettiset muutokset.
MS-tautiin assosioituvia geenejä on tunnistettu noin 200, mutta yksikään potilaista ei varmasti kanna kaikkia mahdollisia MS-tautiin liittyviä geenimuutoksia. Tämä monimuotoisuus selittää sen, miksi MS-tautiin on äärimmäisen vaikeaa löytä parantavaa ja kaikille potilaille soveltuvaa hoitoa.
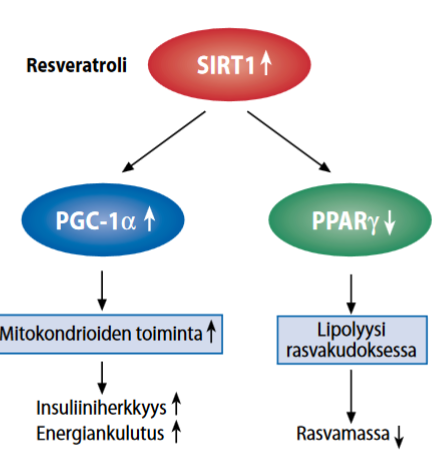
Oksidatiiviseen ainnenvaihduntaan vaikuttavia tekijöitä: PPAR, sirtuiinit ja AMPK
Palataan hapetusreaktioihin, sillä ne vaikuttavat solujen aineenvaihduntaan mm. ravintoaineiden kautta. Oksidatiivinen stressi ja matala-asteinen tulehdus heikentävät elimistön terveyttä ja altistavat kroonisille sairauksille. Immuunivälitteisissä tulehduksellisissa sairauksissa oksidatiivinen stressi ja inflammaatio ylläpitävät ja pahentavat taudin oireita.
Happiradikaalit kaappaavat elektroneja muilta molekyyleiltä
Oksidatiivisella stressillä tarkoitetaan solujen ja laajemmin koko elimistön hapetus-pelkistystilan epätasapainoa. Kun hapettavia tekijöitä on liikaa suhteessa pelkistäviin tekijöihin, oksidatiivinen stressi välittyy reaktiivisten happi- ja typpiradikaalien kautta muihin molekyyleihin.
Reaktiivinen happiradikaali (ROS) on hapesta muodostunut yhdiste, joka sisältää parittoman elektronin. Se pyrkii parilliseen elektronimäärään reagoimalla läheisyydessään olevien muiden yhdisteiden kanssa. Tämä johtaa eräänlaiseen dominoefektiin, jossa happiradikaali vaurioittaa kohtaamansa molekyylin rakennetta ja/tai toimintaa.
Oksidatiivisen metabolismin vaikutusta tehostaa kaksi entsyymiä ja tumareseptori. Entsyymit ovat AMP-aktivoidut proteiinikinaasit: AMPK (Steinberg and Kemp, 2009) sekä sirtuiinit (SIRT), jotka ovat joukko NAD+ -vaikutuksesta aktivoituvia histonideasetylaaseja (Zhang et al., 2011; Rice et al., 2012). Vaikuttava tumareseptori on PPAR-isotyyppi (peroxisome proliferator-activated receptors) Desvergne and Wahli, 1999; Burns and VandenHeuvel, 2007).
Rasvojen energiantuotanto
Keho säilyttää energiaa rasvahappoina, koska rasvahapoissa on hiilihydraatteihin nähden yli kaksinkertainen määrä energiaa painoyksikköä kohden. Rasvahappoja muutetaan energiaksi mitokondrioissa tapahtuvassa beeta-oksidaatiossa:
- Aluksi rasvat hajotetaan rasvahapoiksi ja glyseroliksi. Esimerkiksi triglyseridissä on kolme rasvahappoketjua, jotka ovat kiinnittyneenä glyseroliosaan.
- Glyseroli hapetetaan solulimassa glyseraldehydi-3-fosfaatiksi, joka voidaan käyttää energiantuotantoon (n. 5 % triglyserideistä saatavasta energiasta) tai glukoosin tuottamiseen glukoneogeneesissä. Glukoneogeneesi käyttää glukoosimolekyylin tuottamiseen enemmän energiaa kuin syntyvästä glukoosimolekyylistä vapautuu glykolyysissä ja soluhengityksessä.
- Rasvahapot hapetetaan mitokondrioissa tapahtuvassa beeta-oksidaatiossa (β–oksidaatiossa).
Rasvahapot aktivoidaan edelleen mitokondrion ulkokalvolla kiinnittämällä rasvahapon karboksyyliryhmään koentsyymi A. Näin muodostunut asyyli-KoA kulkee mitokondrion sisäkalvon läpi aktiivisella kuljetuksella. Soluliman asyyli-KoA:lla ja mitokondrion asyyli KoA:lla on eri tehtävät: solulimassa ”rakentava” anabolia ja mitokondriossa ”hajottava” katabolia. - Mitokondrion matriksissa rasvahappo hajotetaan kaksihiilisiksi pätkiksi (asetyyli-KoA).
- Asetyyli KoA (asetyylikoentsyymi A) hapetetaan edelleen sitruunahappokierrossa.
- Elimistön energiantuotannon lopputuotteena syntyy vettä ja hiilidioksidia, jotka poistuvat kehosta mm. hengityksen ja hikoilun kautta.
Mitokondrioissa ja peroksisomeissa tapahtuvaa rasvahappojen beetaoksidaatiota tehostavat PPAR-isotyypit. Beetaoksidaatiossa ravinnon tai kehon varastoimia rasvahappoja käytetään energianlähteenä. PPAR-isotyypit säätelevät beetaoksidaatioon liittyvien geenien transkriptiota ja muodostavat AMPK-sirtuiinipolkuja.
Vähäenerginen ravinto ja liikunta aktivoi AMPK-sirtuiini-PPAR-polun aineenvaihduntaa
AMPK-sirtuiini-PPAR-polku aktivoituu vähäenergisen ravinnon ja fyysisen harjoittelun seurauksena. Aktivaatiota tehostavat kasvisten ja hedelmien sisältämät polyfenolit ja pitkäketjuiset monityydyttämättömät rasvahapot (omega-3). Ligandin aktivoimat PPAR-isotyypit muodostavat kahdesta erilaisesta osasta koostuvia (heterodimeerisiä) komplekseja RXR-reseptorin kanssa.
Käytännössä: Vähäenerginen, omega-3-rasvahappoja ja polyfenoleita sisältävä ravinto tehostaa aineenvaihduntaprosessia, jossa rasvahappoja muutetaan energiaksi beeta-oksidaatiossa.
Vastaavasti runsasenerginen ravinto tehostaa anabolista aineenvaihduntaa ja lipogeneesiä, jossa verenkierrossa olevia sokereita muutetaan varastorasvoiksi. Energiatiheät ravintoaineet edistävät solujen kasvua aktivoimalla SREBP-1c ja SREBP-2 proteiineja (sterol regulatory element-binding proteins), Xu et al., 2013, ja ChREBP (carbohydrate responsive element-binding protein), Xu et al., 2013.
LXR tumareseptorit kontrolloivat SREBP-1c ja SREBP-2-proteiineja, Mitro et al., 2007; Nelissen et al., 2012. Oksysterolit ja glukoosi puolestaan aktivoivat SREBP-1c- ja SREBP-2-proteiineja, jotka osallistuvat lipidien, triglyseridien ja kolesterolin synteesiin.
MS ja inflammaatio: NF-kB ja AP-1)
Ravinnon, inflammaation ja MS-taudin yhteyden kannalta merkityksellisiä ovat kaksi transkriptiotekijää, jotka osallistuvat inflammaatioon ja autoimmuunireaktioihin. Nämä ovat tuman transkriptiotekijä-kB (NF-kB) ja aktivaattoriproteiini (AP-1; Yan and Greer, 2008).
MS-taudissa sekä NF-kB ja AP-1 aktivoituvat vaikuttaen useiden proinflammatoristen geenien ekspressioon ja proinflammatoristen molekyylien tuotantoon. Aktivoitumisen mekanismia ei täysin tunneta, mutta on todennäköistä, että aktivaatioon vaikuttaa virusten, sytokiinien ja oksidatiivisen stressin lisäksi eräät ravintoaineet, kuten tyydyttyneet rasvat, transrasvat.
Tumareseptoreiden aktivaatio
Kaikkien tumareseptoreiden (PPAR, LXR ja VDR) on aktivoiduttava erityisten ligandien avulla. Nämä ligandit voivat olla spesifejä ravintotekijöitä, mikä osoittaa, kuinka solut reagoivat ravintoaineisiin ja säätelevät energian homeostaasia. Samalla tämä mekanismi on kuin molekylaarinen avain, joka auttaa ymmärtämään kuinka ravintoaineet vaikuttavat tulehduksellisten sairauksien etenemiseen (Heneka et al., 2007; Zhang-Gandhi and Drew, 2007; Krishnan and Feldman, 2010; Cui et al., 2011; Schnegg and Robbins, 2011; Gray et al., 2012).
”Therefore, each of the three nuclear receptors—PPAR, LXR, and VDR—competes for the binding to RA-RXR and forms hetero-complexes that can inhibit NF-kB and exert a tight control over the expression of inflammatory genes, thus integrating metabolic and inflammatory signaling. It is clear that there is competition between the three receptors PPAR, LXR, and VDR-D, for the binding with RA-RXR, but this competition should have an influence only on metabolism and not on inflammation, because it is not yet known which of the three heterodimers is more effective in inhibiting NF-kB.”
Proinflammatoristen molekyylien tuotanto MS-taudin pahenemisvaiheen aikana on biosynteettinen prosessi, jota ylläpitää ja pahentaa runsasenerginen ruokavalio. Toisaalta inflammaatioon assosioituvan relapsin oireita ja kestoa voi helpottaa vähäenergisellä ruokavaliolla.
”In principle, what favors anabolism will promote the inflammatory processes, while what favors catabolism will contrast them.”

Kuvan lähde:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4342365/figure/fig4-1759091414568185/
Tästä artikkelisarjasta on tullut sellainen iisakin kirkko, joka ei näytä koskaan valmistuvan. Aihe on älyttömän kiinnostava.