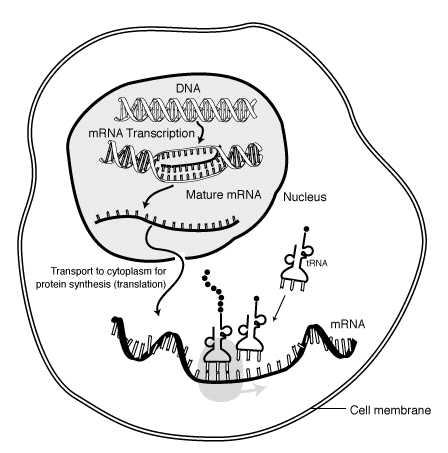
Lähetti-RNA (mRNA) ja mRNA-rokotteet
SARS-CoV-2-rokotteiden sisältämä piikkiproteiinia koodaava mRNA herättää monissa huolta. Se on uusi ja useimmille tuntematon rokotusteknologia, josta liikkuu paljon misinformaatiota. Kuinka pitkään lähetti-RNA toimii ja voiko mRNA ohjata piikkiproteiinien valmistamista solun kuolemaan asti tai solunjakautumisen jälkeen uusissa soluissa? Onko rokote COVID-19-infektiota vaarallisempi?
Halusin tietää enemmän, joten kokosin mRNA:sta ja siihen perustuvasta rokoteteknologiasta tämän tietopaketin.
Saatteeksi
Rokotukset ovat pitkään kuuluneet olennaisena osana kansanterveysohjelmiin ympäri maailman. Suomeen isorokkorokotukset tulivat jo 1800-luvun alussa. Monet rokotukset olivat pakollisia 1950-luvulle asti. Rokotusohjelmat ovat vähentäneet yleisvaarallisten tartuntatautien leviämistä ja vakavuutta. Erityisesti kanslliset rokotukset ovat vähentäneet lapsikuolleisuutta. Rokotusstrategioiden menestys lasten suojelemiseksi poliolta, hepatiitti B:ltä, tuhkarokolta, influenssalta ja pneumokokkiepidemioilta on vahvasti osoitettu.
COVID-19-pandemia loi kiireellisen tarpeen tehokkaalle rokotteelle. Koronarokoteohjelmassa hyödynnettiin ähetti-RNA-tekniikkaa, joka luokitellaan uuden sukupolven rokotusteknologiaksi.
Lääketiede ja -teknologia ovat edistyneet jättiläisen harppauksin viime vuosisadalta mm. parempien ja tarkempien tutkimusvälineiden ja tehokkaampien tietokoneiden ansiosta. Koronarokotteiden kehitystä nopeutti 30 vuoden tutkimustyö mRNA-menetelmästä, tutkimusryhmien rajat ylittävä yhteistyö sekä SARS- ja MERS-epidemioiden kesken jääneiden rokoteohjelmien kokoama arvokas data.
Synteettisten mRNA-alustojen vuosikymmenien tutkimus mm. syöpien hoitomuotona ja rokotteena malariaa, rabiesta, ebolaa, AIDSia, influenssaa ja muita tartuntatauteja vastaan osoittautui arvokkaaksi, kun sekä Modernan että Pfizer/BioNTechin COVID-19-mRNA-rokotteet saivat hätäkäyttöluvan.
COVID-19-rokotteiden myötä mRNA-teknologiat ovat nousseet jokaisen tietoon, mutta menetelmä tunnetaan vielä huonosti. Se aiheuttaa pelkoja.
Johdanto: lähetti- RNA:n transkriptio ja translaatio
mRNA-molekyylin ”elinkaari” eukaryoottisolussa vaihtelee minuuteista päiviin, mutta yleensä mRNA hajoaa solussa varsin nopeasti. Niin myös koronarokotteen lihassoluihin kuljettama mRNA-molekyyli, joka hajoaa parissa päivässä.
Transkriptio tapahtuu, kun RNA kopioidaan DNA:sta. Transkription aikana RNA-polymeraasi tekee tarvittaessa kopion DNA:n geenistä mRNA:han. Prosessi eroaa hieman eukaryooteilla ja prokaryooteilla. Yksi huomattava ero on, että prokaryoottinen RNA-polymeraasi assosioituu DNA:ta prosessoivien entsyymien kanssa transkription aikana, jotta prosessointi voi jatkua transkription aikana. Tämä tekee uudesta mRNA:sta kaksijuosteisen tuottamalla komplementaarisen juosteen (tRNA-juoste), joka ei pysty koodaamaan rakenteita emäspariutumisesta.
mRNA:n templaatti on tRNA:n komplementaarinen juoste, joka on sekvenssiltään identtinen sen antikodonisekvenssin kanssa, johon DNA sitoutuu.
Lyhytikäistä, prosessoimatonta tai osittain prosessoitua molekyyliä kutsutaan prekursori-mRNA:ksi tai pre-mRNA:ksi; kun se on täysin prosessoitu, sitä kutsutaan kypsäksi mRNA:ksi.
Normaalisti mRNA transkriptoidaan solun tumassa. Transkriptiossa RNA-polymeraasi kopioi DNA:ssa olevaa geneettistä koodia RNA:ksi. Transkriptio on proteiinisynteesin ensimmäinen vaihe, mutta transkriptiossa syntyy myös t-RNA-, r-RNA- ja s-RNA-molekyylejä.
Transkriptiossa syntynyt mRNA-molekyyli kuljetetaan tumasta solun sytoplasmaan, jossa solun miljoonat soluorganellit – ribosomit tulkitsevat lähetti-RNA:n. Translaatiossa lähetti-RNA tulkitaan ja sen ohjeen perusteella rakennetaan sitä vastaava aminohappoketju, eli proteiini.
Translaatiossa lähetti-RNA:n nukleotidien järjestys käännetään geneettisen koodin mukaisesti polypeptidiketjun aminohappojärjestykseksi. Tätä varten lähetti-RNA kuljetetaan solulimassa sijaitsevan ribosomin luo. Jos tuotettava proteiini on kalvoproteiini, solun sisäisen kalvojärjestelmän osa tai solusta ulos eritettävä proteiini kuten hormoni, ribosomi kiinnittyy solulimakalvostoon tai tumakotelon ulkoseinään. Muuten proteiinisynteesi jatkuu vapaana solulimassa olevassa ribosomissa.
Ribosomin pinnalla siirtäjä-RNA-molekyylien paikalle kuljettamat aminohapot liitetään toisiinsa peptidisidoksilla pitkäksi ketjuksi, jonka järjestyksen määrää lähetti-RNA:n emäsjärjestys. Tämä perustuu kolmen peräkkäisen emäksen ryhmiin, kodoneihin, joita kutakin vastaa tietty aminohappo. Proteiinisynteesin aloituskohtaa merkitsee oma kodoni, joka koodaa myös metioniini-aminohapon liittämistä ketjuun. Ribosomi lopettaa polypeptidiketjun muodostamisen kohdattuaan yhden kolmesta lopetuskodonista.
Lopuksi mRNA-molekyyli hajoaa.
Lähetti-RNA (mRNA)
Nisäkässolujen kolme tärkeintä RNA-tyyppiä ovat lähetti-RNA, ribosomaalinen RNA (rRNA) ja siirtäjä-RNA (tRNA).
Ribonukleiinihappo (RNA) on luonnollinen molekyyli, joka löytyy kaikista soluista. RNA:ta on monia tyyppejä, joista jokaisella on oma tehtävänsä molekyylibiologiassa. mRNA toimii tärkeänä viestimolekyylinä soluissa.
Lähetti-RNA-molekyyli kuljettaa proteiinien rakennusohjeet tumasta sytoplasmaan. mRNA:n aminohapposekvenssi kertoo koska ja millaisia proteiineja solun on tarkoitus valmistaa. mRNA:n koodi kopioidaan solun tumassa olevasta DNA-juosteesta prosessissa, jota kutsutaan transkriptioksi.
mRNA kuljetetaan sytoplasmaan (solun sisältämä liuos), jossa solun proteiinintuotantokoneisto lukee ja kääntää koodin. Tuloksena on proteiini: entsyymi, vasta-aine, hormoni tai solun rakennekomponentti.
mRNA on yksijuosteinen ribonukleiinihappo-molekyyli, joka vastaa geenin geneettistä sekvenssiä. Ribosomit lukevat mRNA-molekyylin proteiinisynteesissä. Transkriptioprosessin aikana RNA-polymeraasi-entsyymi muuntaa geenin primaariseksi transkriptio-mRNA:ksi, joka tunnetaan myös nimellä pre-mRNA.
Tämä pre-mRNA sisältää usein proteiineja koodaamattomia introneita. Intronit voivat kuitenkin sisältää alueita, jotka säätelevät proteiinisynteesiä. Intronit poistetaan RNA:n silmukointiprosessissa, jolloin jäljelle jää vain eksonit, eli alueet, jotka koodaavat proteiinia. Tämä eksonisekvenssi muodostaa valmiin mRNA:n.
Ribosomit lukevat valmiin mRNA:n, ja valmistavat mRNA:n aminohapposekvenssin mukaisen proteiinin käyttämällä siirto-RNA:n (tRNA) kuljettamia aminohappoja. Tämä prosessi tunnetaan translaationa.
RNA:ssa ja DNA:ssa geneettinen informaatio sisältyy nukleotidisekvensseihin, jotka on järjestetty kodoneiksi. Kussakin kodonissa on kolme ribonukleotidiä. Jokainen kodoni koodaa tiettyä aminohappoa. Poikkeuksena on proteiinisynteesin lopettavat lopetuskodonit, jotka eivät koodaa aminohappoja.
Kodonien translaatio aminohappoiksi vaatii kahta muuta RNA-tyyppiä:
1. siirtäjä-RNA (tRNA) tunnistaa kodonin ja tuo proteiinisynteesiin kodonia vastaavan aminohapon
2. ribosomaalinen RNA (rRNA) on ribosomin proteiininvalmistuskoneiston keskeinen komponentti.
mRNA:n historia
Sydney Brenner ja Francis Crick saivat ensimmäisenä idean mRNA:sta 15. huhtikuuta 1960 King’s Collegessa, Cambridgessa, kun François Jacob kertoi heille kokeesta, jonka tekivät Arthur Pardee, hän itse ja Jacques Monod [1].
Crickin rohkaisemana Brenner ja Jacob ryhtyivät testaamaan uutta hypoteesia, ja he ottivat yhteyttä Matthew Meselsoniin Kalifornian teknologiainstituutissa [1]. Kesällä 1960 Brenner, Jacob ja Meselson suorittivat kokeen Meselsonin laboratoriossa Caltechissa. Tämä vahvisti mRNA:n olemassaolon [1].
Samana syksynä Jacob ja Monod loivat nimen ”lähetti-RNA” ja kehittivät ensimmäisen teoreettisen kehyksen selittämään sen toimintaa [1]. Helmikuussa 1961 James Watson kertoi, että hänen tutkimusryhmänsä teki samanlaisia tutkimuksia; Brenner ja muut suostuivat Watsonin pyyntöön lykätä tutkimustulosten julkaisemista [1]. Tämän seurauksena Brennerin ja Watsonin artikkelit julkaistiin samanaikaisesti samassa Nature-lehden numerossa toukokuussa 1961, kun taas samassa kuussa Jacob ja Monod julkaisivat teoreettisen viitekehyksensä mRNA:lle Journal of Molecular Biology -lehdessä [1].
Synteesi, prosessointi ja toiminta
Noin 40 vuotta sitten tutkijat havaitsivat, että he pystyivät jäljittelemään luonnollista transkriptiota ja tuottamaan synteettistä mRNA:ta ilman solua. Prosessi, joka tunnetaan nimellä in vitro -transkriptio, voi tuottaa mRNA-molekyylejä koeputkessa olevasta DNA-juosteesta.
Tämä edellyttää RNA-polymeraasi-entsyymiä ja nukleotideja (molekyylejä, jotka ovat DNA:n ja RNA:n rakennuspalikoita. Kun polymeraasi ja nukleotidit sekoitetaan, polymeraasi-entsyymi lukee DNA-juosteen ja muuntaa koodin mRNA-juosteeksi yhdistämällä eri nukleotidit yhteen oikeassa järjestyksessä.
Kun in vitro -transkriptoitu mRNA viedään soluun, solun proteiinintuotantokoneisto ”lukee” sen samalla tavalla kuin luonnollisen mRNA:n. Menetelmää voidaan käyttää synteettisen mRNA:n tuottamiseen, joka koodaa mitä tahansa kiinnostavaa proteiinia.
Rokotteiden mRNA koodaa osaa virusproteiinista, joka tunnetaan antigeeninä. Kun antigeeni on käännetty, se aktivoi immuunivasteen, joka auttaa antamaan suojan virukselta.
mRNA on hyvin lyhytikäinen, eikä se pysty muuttamaan solun DNA:ta. Siten se on turvallinen menetelmä.
Synteettinen mRNA on turvallista rokotteissa ja syöpähoidoissa. In vitro -transkription etu on, että se ei vaadi soluja tuottamaan mRNA:ta. Tällä on valmistusetuja muihin rokoteteknologioihin verrattuna – esimerkiksi nopeus ja pienemmät biologiset turvallisuusriskit.
Kesti vain 25 päivää valmistaa erä Modernan lipidinanohiukkas-mRNA-rokotetta kliinisiin kokeisiin.Siitä tuli maaliskuussa 2020 ensimmäinen COVID-19-rokote, joka eteni kliinisiin ihmiskokeisiin.
Koska in vitro -transkriptio on soluton, synteettisten mRNA-molekyylien valmistusmenetelmä on joustava. Uusia rokotteita tai hoitoja voidaan suunnitella olemassaolevien menetelmien pohjalta. DNA-koodin korvaamalla tutkimuslaitokset voivat helposti siirtyä yhden mRNA-rokotteen valmistamisesta toiseen. Tämä voi osoittautua elintärkeäksi nopeille rokotevasteille tulevissa epidemioissa ja pandemioissa.
mRNA-molekyylin lyhyt olemassaoloaika alkaa transkriptiosta ja päättyy lopulta hajoamiseen. mRNA-molekyyliä voidaan käsitellä, muokata ja kuljettaa ennen translaatiota. Eukaryoottiset mRNA-molekyylit vaativat usein laajaa käsittelyä ja kuljetusta, kun taas prokaryoottiset mRNA-molekyylit eivät. Eukaryoottisen mRNA:n molekyyliä ja sitä ympäröiviä proteiineja kutsutaan yhdessä lähetti-RNP:ksi.
mRNA rokotteissa
Lähetti-RNA:n tunnetuin tehtävä on geneettisen tiedon välittäminen DNA:sta proteiinien aminohappojärjestykseksi. Proteiinin valmistusohjeet siirtyvät lähetti-RNA:n muodossa proteiinitehtaisiin, ribosomeihin, joiden keskeisiä rakenneosia ovat ribosomaalisen RNA:n alayksiköt. Aminohappojen toimittajina ovat pienet siirtäjä-RNA-molekyylit.
Käytössä olevat mRNA-rokotteet ovat hyötyneet vuosien tutkimuksesta, suunnittelusta ja optimoinnista. Tieto siitä, kuinka synteettinen RNA tunnistetaan soluissa, on osoittautunut välttämättömäksi tehokkaiden rokotteiden kehittämisessä.
Tyypillisesti mRNA koodaa tunnettua virusantigeeniä. COVID-19-mRNA-rokotteiden tapauksessa on käytetty SARS-CoV-2-piikkiproteiinia tai reseptoriin sitoutuvaa domeenia koodaavia sekvenssejä.
Nämä antigeeniä koodaavat mRNA-molekyylit liitetään hyvin pieniin partikkeleihin, jotka koostuvat pääasiassa lipideistä (rasvoista). Lipidipartikkelilla on kaksi päätehtävää: se suojaa mRNA:ta hajoamiselta ja auttaa kuljettamaan rokotteen soluun.
Solun sytoplasmassa rokotteen sisältämä mRNA transloituu antigeeniksi, joka käynnistää immuunivasteen. Tämä prosessi aktivoi immuunijärjestelmän kehittämään menetelmiä antigeenin neutraloimiseksi. Immuunijärjestelmän primaarivaste kestää yleensä muutaman viikon, ennen kuin hankinnainen (adaptiivinen) immuunijärjestelmä kypsyy ja synkronoituu uhkaan tuottaen toimivan immuunivasteen.
mRNA-rokotteiden on osoitettu stimuloivan adaptiivisen immuunivajärjestelmän molempia käsivarsia: B-solujen tuottama vasta-ainevälitteinen ja T-solujen tuottama soluvälitteinen immuniteetti ovat tärkeitä tehokkaan immuunivasteen muodostamisen kannalta. Humoraalinen eli vasta-ainevälitteinen immuniteetti tuottaa antigeenille herkistyneitä immunoglobuliineja ja soluvälitteinen immuniteetti havaitsee ja tuhoaa infektoituneita soluja.
B-solujen tuottamat immunoglobuliinit jaetaan viiteen ryhmään: IgA, IgD, IgE, IgG ja IgM ja ne voivat tuottaa arviolta 1012 erilaista antigeeniin kiinnittyvää reseptoria. Käytännössä B-solut voivat tuottaa rajattomasti erilaisia vasta-aineita.
Y:n muotoiset immunoglobuliinit sitoutuvat antigeeniin, eli piikkiproteiiniin. SARS-CoV-2 tarvitsee piikkiproteiinin telakoituakseen isäntäorganismin kohdesolun ACE2-reseptoriin, joten vasta-aineet estävät tehokkaasti viruksen kyvyn infektoida soluja. Tämä on tärkeää infektion hillinnän kannalta. Virukset voivat lisääntyä vain kaappaamissaan soluissa, joten vasta-aineet estävät tehokkaasti viruksen lisääntymisen. Vasta-aineet toimivat myös tunnisteina immuunijärjestelmän syöjäsoluille, eli fagosyyteille, kuten makrofageille, jotka syövät immunoglobuliinien merkitsemät virukset.
Käytössä olevat mRNA COVID-19 -rokoteohjelmat käyttävät kahden annoksen (prime-boost) -lähestymistapaa, jonka tavoitteena on vahvistaa adaptiivista immuunivastetta SARS-CoV-2-virusta vastaan. B-solujen ja niiden tuottamien immunoglobuliinien määrä vähenee muutamissa kuukausissa. Tämä heikentää akuuttia humoraalista immuunivastetta: immuunijärjestelmä siirtyy akuutista vasta-ainevälitteisestä ”hälytystilasta” immunologisen muistin ylläpitämään ”valmiustilaan”.
Immunologisen muistin aktivoima immuunivaste tuottaa uusia B- ja T-soluja, mutta tämän immuunivasteen kehittyminen vie pari päivää. Immunologiseen muistiin perustuva immuniteetti ei ole yhtä nopea kuin rokotusten tai sairastetun infektion tuottama akuutti vasta-ainevälitteinen immuniteetti. Tämän vuoksi monissa maissa on jo otettu käyttöön 3. rokoteannos, joka vahvistaa vasta-ainevälitteistä immuunipuolustusta.
mRNA-rokotetyyppi, jota kutsutaan itsevahvistuvaksi RNA:ksi (self-amplifying), saattaa edellyttää vain yhden rokoteannoksen saman suojatason saavuttamiseksi kuin nyt käytettävien rokotteiden 2-3 annosta. Solussa nämä itsestään monistuvat RNA-rokotteet voivat kopioida mRNA-koodin.
Tämä tarkoittaa, että pienemmällä mRNA:lla voidaan tuottaa enemmän antigeeniä. Useat tällä hetkellä kliinisissä tutkimuksissa olevissa COVID-19-RNA-rokotteissa tutkivat itseään vahvistavia RNA-tekniikoita.
mRNA ja itseään monistava RNA osoittavat potentiaalia monien tartuntatautien, kuten influenssan, hengitysteiden synsyyttiviruksen, rabieksen, ebolan, malarian ja HIV-1:n rokotemekanismina. Yhdessä terapeuttisten sovellusten kanssa, etenkin syöpien hoidon immunoterapiana, mRNA-teknologiat paranevat ja laajenevat edelleen, ja ne ovat olennainen osa uuden sukupolven lääkkeitä.
Aitotumaisten pre-mRNA-molekyylin prosessointi
mRNA:n prosessointi vaihtelee suuresti aitotumaisten (eykaryoottien), bakteerien ja arkkien välillä. Ei-eukaryoottinen mRNA on pohjimmiltaan kypsä transkription jälkeen eikä vaadi käsittelyä, paitsi harvoissa tapauksissa [2]. Eukaryoottinen pre-mRNA vaatii kuitenkin useita prosessointivaiheita ennen sen kuljettamista sytoplasmaan ja sen translaatiota ribosomin toimesta.
Aitotumaisten soluissa transkriptiossa syntyy lähetti-RNA:n esiastetta pre-mRNA:ta, jota muokataan kolmella tavalla ennen kuin se pääsee kulkemaan tumasta solulimaan. Ensimmäisenä pre-mRNA:n 5′-päähän liitetään GTP, johon on liittynyt metyyliryhmä. Toiseksi pre-mRNA:n 3′-päähän liitetään Poly(A)-häntä, joka koostuu pelkästään adenosiinimonofosfaateista eli adeniinin sisältävistä RNA-nukleotideista. Päiden muokkaus suojaa lähetti-RNA:ta sitä pilkkovilta entsyymeiltä ja auttaa solua tunnistamaan lähetti-RNA:n. Lopuksi intronit poistetaan silmukoinnilla ennen lähetti-RNA:n poistumista tumasta.
Esitumallisten soluissa ei ole tumaa eikä niiden DNA:ssa ole introneita, joten solulimaan syntyvää lähetti-RNA:ta ei tarvitse muokata, vaan sitä voidaan käyttää translaatiossa sellaisenaan. Lisäksi esitumallisten soluissa transkriptio ja translaatio voivat tapahtua samanaikaisesti.
Ei-koodaavat alueet eli UTR:t (engl. Un-translated regions) ovat lähetti-RNA:ssa ennen aloituskodonia ja lopetuskodonin jälkeen esiintyviä sekvenssejä, jotka eivät translaatiossa koodaa mitään proteiinia.
Alueita kutsutaan myös 5′-UTR:ksi ja 3′-UTR:ksi. Ne kopioidaan transkriptiossa samaan tapaan kuin koodaavat alueetkin ja ne kuuluvat eksoneihin, minkä takia niitä ei poisteta pre-mRNA:ta muokattaessa. Ei-koodaavilla alueilla voidaan katsoa olevan useita rooleja geenien ilmentymisessä, esimerkiksi lähetti-RNA:n vakautus, paikallistaminen ja translaation tehostaminen. Ei-koodaavan alueen kyky toimia riippuu kuitenkin sekvenssistä ja voi vaihdella eri lähetti-RNA:issa.
Ei-koodaavat alueet voivat huolehtia translaation tehokkuudesta tai joissain tapauksissa kokonaan estää sen. Ei-koodaaviin alueisiin sitoutuvat proteiinit vaikuttavat ehkä translaatioon vaikuttamalla ribosomin kykyyn sitoutua lähetti-RNA:han. 3′-UTR:iin sitoutuva mikroRNA saattaa myös vaikuttaa translaation tehokkuuteen tai lähetti-RNA:n vakauteen.
3′-UTR vastaa todennäköisesti lähetti-RNA:n sijainnista solulimassa. Jos tietyllä solun alueella tarvitaan proteiinia, sitä voidaan tuottaa kyseisellä alueella. Tällaisia tapauksia varten 3′-UTR:ssä voi olla sekvenssejä, jotka sallivat lähetti-RNA:n siirtyä tällaiselle alueelle translaatiota varten.
Jotkin ei-koodaavilla alueilla olevat elementit muodostavat sekundäärirakenteen, kun geeni kopioidaan transkriptiossa RNA:ksi. Niillä on myös osansa lähetti-RNA:n säätelyssä. Jotkin niistä ovat proteiinien sitoutumispaikkoja. Ribokytkimiin voi sitoutua pieniä molekyylejä, jolloin lähetti-RNA:n koodaaman proteiinin tuotannossa tapahtuu muutos. Tällaisissa tapauksissa lähetti-RNA säätelee itse itseään.
Poly(A)-häntä on pre-mRNA:n 3′-päähän lisättävä, usein satoja adeniininukleotideja sisältävä sekvenssi. Häntä estää lähetti-RNA:ta hajoamasta.
Lähetti-RNA:ta, jossa on vain yhden proteiinin translaatioon tarvittava ohje, kutsutaan monokistriseksi. Useimpien aitotumaisten solujen lähetti-RNA on monokistrista.Sitä vastoin esitumallisten solujen lähetti-RNA on polykistrista, eli siinä on useita avoimia lukukehyksiä eli proteiinin translaatioon tarvittavia ohjeita.
Yleensä polykistrisen lähetti-RNA:n koodaamilla proteiineilla on jokin yhteinen ominaisuus – ne voivat esimerkiksi muodostaa yhdessä proteiinikompleksin – ja niitä koodaavia sekvenssejä säädellään yhteisellä säätelyalueella, jossa on promoottori ja operaattori. Myös bakteerien ja arkkien lähetti-RNA ja ihmisen mitokondriaalinen lähetti-RNA on suurimmaksi osaksi polykistristä. Di- tai bikistriseksi kutsutaan lähetti-RNA:ta, joka koodaa kahta proteiinia.
Aitotumaisten soluissa lähetti-RNA-molekyylit muodostavat pyöreitä rakenteita eIF4E-proteiinin ja poly(A):n sitojaproteiinin välisten vuorovaikutuksien takia. Molemmat proteiinit sitoutuvat eIF4G-proteiiniin ja muodostavat siten mRNA-proteiini-mRNA-sillan. Rengastumisen uskotaan tukevan ribosomien kiertoa ja tehostavan translaatiota, ja ehkä myös toimivan varmistimena sille, että vain ehjät lähetti-RNA-molekyylit osallistuvat translaatioon.[8] Ehjissä lähetti-RNA-molekyyleissä on poly(A)-häntä ja 5′-cap.
Silmukointi
RNA-silmukointi on eukaryoottisen pre-mRNA:n prosessi, jonka seurauksena on kypsä mRNA. Silmukoinnissa (splicing) intronit tai outronit (ei-koodaavat alueet) poistetaan ja eksonit (koodaavat alueet) liitetään yhteen.
Silmukointi on geenin transkription (DNA:sta esi-mRNA:ksi) ja translaation (mRNA:sta proteiiniksi) välissä tapahtuva prosessi. Silmukoinnissa geenistä poistetaan transkriptoidussa esi-mRNA:ssa olevat jaksot, jotka eivät vaikuta proteiinin sekvenssiin (intronit), jolloin jäljelle jäävät vain geenin koodaavat osat eli eksonit.
Eksoneiden, joiden sekvenssi määrää proteiinien rakenteen, välissä on introneita, jotka poistetaan ennen proteiinisynteesiä. Proteiinisynteesi alkaa kuitenkin koko geenin – eksoneiden ja introneiden – transkriptiolla esi-mRNA:ksi.
Esi-mRNA:sta poistetaan tumassa intronit, jolloin syntyy valmiita mRNA-juosteita (lähetti-RNA:ta), jotka siirretään tumasta solulimaan proteiinisynteesiä varten.
Silmukointikohdan tunnistus perustuu introneiden päissä oleviin konservoituneisiin jaksoihin. RNA:n 5′-päässä ovat nukleotidit GU ja 3′-päässä AG. Silmukointireaktiossa esi-mRNA katkeaa ensin intronin 5′-päästä, kun intronissa oleva adeniinin -OH-ryhmä hyökkää intronin 5′-pään fosforisidokseen. Molekyyli muodostaa tällöin lasson (engl. lariat). Toisessa vaiheessa 5′-päässä ollut eksoni hyökkää 3′-pään eksonin fosfaattiryhmään.
Esi-mRNA:n silmukointi voi tapahtua vain yhdellä tavalla, mutta usein samasta geenistä voidaan silmukoinnin avulla luoda erilaisia mRNA-molekyylejä ja siten erilaisia proteiineja. Tämä mahdollistaa eliöiden kasvavan monimutkaisuuden ja nopeamman evoluution. Vaikka silmukoinnin lopputuotteet voivat olla erilaisia, eksonit ovat niissä aina samassa järjestyksessä kuin esi-mRNA:ssa.
Ehkä tärkein viimeaikainen havainto, joka korostaa lähetti-RNA-eli mRNA-molekyylien itsenäisiä tehtäviä, liittyy RNA-tuotannon välivaiheena esiintyvän esiaste-RNA:n muokkauksen keskeisimpään tapahtumaan, intronien poistoon eli silmukointiin.
Lukuisten proteiinien rakenne muuntuu kehityksen eri vaiheissa tai kudosspesifisesti vaihtoehtoisen silmukoinnin avulla, jolloin muodostuvan valkuaisaineen rakenne vaihtelee sen mukaan, minkä eksonin informaatio säilyy tai poistuu esiaste-RNA:sta (Beardsley 1991). Esimerkiksi transkriptiota säätelevien proteiinien esiaste-RNA:itten vaihtoehtoinen silmukointi vaikuttaa oleellisesti niiden biologiseen aktiivisuuteen ja sitä kautta moniin solun tapahtumiin (Chambon 1981). Itse vaihtoehtoisen silmukoinnin säätely tunnetaan huonosti.
5′ huppurakenteen (cap) lisäys
Esiaste-RNA:n muokkaaminen mRNA:ksi alkaa tumassa heti transkription jälkeen. Lähetti-RNA:n alkuun, 5–päähän, liittyy erityinen huppurakenne (cap) ja loppuun, 3–päähän, 100–200 adenyyliä sisältävä poly-A-häntä, jota tarvitaan sekä mRNA:n siirtoon että stabilointiin.
5′ cap (kutsutaan myös RNA capiksi, RNA 7-metyyliguanosiini cap tai RNA m7G cap) on modifioitu guaniininukleotidi, joka on lisätty aitotumallisen lähetti-RNA:n ”etu-” tai 5′-päähän pian transkription alkamisen jälkeen.
5′-huppurakenne koostuu terminaalisesta 7-metyyliguanosiinitähteestä, joka on liitetty 5′-5′-trifosfaattisidoksella ensimmäiseen transkriptoituun nukleotidiin. Sen läsnäolo on kriittinen ribosomin tunnistamiselle ja suojautumiselle RNaaseilta.
Huppurakenteen lisäys (capping) on kytketty transkriptioon ja tapahtuu kotranskriptionaalisesti siten, että kumpikin vaikuttaa toisiinsa. Pian transkription alkamisen jälkeen syntetisoitavan mRNA:n 5′-pää sitoutuu RNA-polymeraasiin liittyvään cap-syntetisoivaan kompleksiin. Tämä entsymaattinen kompleksi katalysoi kemiallisia reaktioita, joita tarvitaan mRNA:n sulkemiseen. Synteesi etenee monivaiheisena biokemiallisena reaktiona.
Editointi
Joissakin tapauksissa mRNA:ta muokataan, mikä muuttaa kyseisen mRNA:n nukleotidikoostumusta. Esimerkki: on apolipoproteiini B:n mRNA, jota muokataan joissakin kudoksissa, mutta ei toisissa. Muokkaus luo varhaisen lopetuskodonin, joka translaation jälkeen tuottaa lyhyemmän proteiinin.
Polyadenylaatio
Polyadenylaatio on polyadenylyyliosan kovalenttinen kytkeminen lähetti-RNA-molekyyliin. Aitotumallisissa organismeissa useimmat lähetti-RNA (mRNA) -molekyylit ovat polyadenyloituneita 3′-päästä, mutta viimeaikaiset tutkimukset ovat osoittaneet, että lyhyet uridiinijaksot (oligouridylaatio) ovat myös yleisiä[3].
Poly(A)-häntä ja siihen sitoutunut proteiini auttavat suojaamaan mRNA:ta eksonukleaasien aiheuttamalta hajoamiselta.
Polyadenylaatio on tärkeä myös transkription lopettamiselle, mRNA:n viennille tumasta solulimaan ja translaatiolle. mRNA voidaan myös polyadenyloida prokaryoottisissa organismeissa, joissa poly(A)-hännät helpottavat, mutta eivät estä eksonukleolyyttistä hajoamista.
Polyadenylaatio tapahtuu transkription aikana ja/tai välittömästi sen jälkeen. Kun transkriptio on lopetettu, mRNA-ketju katkaistaan RNA-polymeraasiin liittyvän endonukleaasikompleksin vaikutuksesta. Kun mRNA on pilkottu, noin 250 adenosiinitähdettä lisätään vapaaseen 3′-päähän katkaisukohtaan. Tätä reaktiota katalysoi polyadenylaattipolymeraasi.
Aivan kuten vaihtoehtoisessa silmukointissa, mRNA:ssa voi olla useampi kuin yksi polyadenylaatiovariantti. Myös polyadenylaatiokohdan mutaatioita esiintyy. Geenin primaarinen RNA-transkripti katkaistaan poly-A-lisäyskohdasta ja RNA:n 3′-päähän lisätään 100–200 A:ta. Jos tätä kohtaa muutetaan, muodostuu epänormaalin pitkä ja epästabiili mRNA-konstrukti.
Kuljettaminen
Toinen ero eukaryoottien ja prokaryoottien välillä on mRNA:n kuljetus. Koska eukaryoottinen transkriptio ja translaatio ovat erillään toisistaan, eukaryoottisia mRNA:ita on kuljetettava tumasta sytoplasmaan. Prosessia voidaan säädellä erilaisilla signalointireiteillä [4].
Kypsät mRNA:t tunnistetaan niiden prosessoitujen modifikaatioiden perusteella ja viedään sitten tuman huokosten läpi sitoutumalla cap-sitoviin proteiineihin CBP20 ja CBP80 [5], sekä transkriptio/vientikompleksiin (TREX) [6][7].
Eukaryoottisoluilla on tunnistettu useita mRNA:n vientireittejä [8]. Tilallisesti monimutkaisissa soluissa jotkin mRNA:t kuljetetaan tiettyihin subsellulaarisiin kohteisiin. Kypsissä neuroneissa tietty mRNA kuljetetaan somasta dendriitteihin.
Yksi mRNA:n translaatiokohta on polyribosomeissa, jotka ovat selektiivisesti lokalisoituneet synapsien alle [9]. Arc/Arg3.1:n mRNA:n indusoi synaptinen aktiivisuus ja se lokalisoituu selektiivisesti aktiivisten synapsien lähelle NMDA-reseptorien tuottamien signaalien perusteella[10].
Myös muut mRNA:t siirtyvät dendriitteihin vasteena ulkoisille ärsykkeille, kuten β-aktiini-mRNA:lle [11]. Tumasta viedään aktiinin mRNA assosioituu ZBP1:een ja 40S-alayksikköön. Kompleksi sitoutuu motoriseen proteiiniin ja kuljetetaan kohdepaikkaan (neuriittijatke) solurunkoa pitkin. Lopulta Src fosforyloi ZBP1:n kääntämisen aloittamiseksi [12]. Kehittyvissä neuroneissa mRNA:t kuljetetaan myös kasvaviin aksoneihin ja erityisesti kasvukartioihin. Monet mRNA:t on merkitty niin sanotuilla ”postinumeroilla”, jotka kohdistavat niiden kuljetuksen tiettyyn paikkaan [13].
Translaatio
Koska prokaryoottista mRNA:ta ei tarvitse käsitellä tai kuljettaa, ribosomin suorittama translaatio voi alkaa välittömästi transkription päättymisen jälkeen.
Siksi voidaan sanoa, että prokaryoottinen translaatio on kytketty transkriptioon ja tapahtuu ko-transkriptionaalisesti. Eukaryoottinen mRNA, joka on prosessoitu ja kuljetettu sytoplasmaan (eli kypsä mRNA), voidaan sitten kääntää ribosomin toimesta.
Translaatio voi tapahtua ribosomeissa, jotka kelluvat vapaasti sytoplasmassa tai jotka signaalintunnistuspartikkeli ohjaa endoplasmiseen retikulumiin. Siksi, toisin kuin prokaryooteissa, eukaryoottinen translaatio ei ole suoraan kytketty transkriptioon. Joissakin yhteyksissä on jopa mahdollista, että alentuneisiin mRNA-tasoihin liittyy kohonneita proteiinitasoja, kuten on havaittu EEF1A1:n mRNA/proteiinitasoilla rintasyövässä [14].
Rakenne
Kypsän eukaryoottisen mRNA:n rakenne.
Täysin prosessoitu mRNA sisältää 5′ capin, 5′ UTR:n, koodaavan alueen, 3′ UTR:n ja poly(A) hännän.
Koodaavat alueet
Koodaavat alueet koostuvat kodoneista, jotka ribosomi dekoodaa ja muuntaa proteiineiksi; eukaryooteissa yleensä yhdeksi ja prokaryooteiksi useiksi. Koodaavat alueet alkavat aloituskodonilla ja päättyvät lopetuskodoniin.
Yleensä aloituskodoni on AUG-tripletti ja lopetuskodoni on UAG (”keltainen”), UAA (”okra”) tai UGA (”opaali”). Koodaavia alueita yleensä stabiloivat sisäiset emäsparit, mikä estää hajoamisen [15][16]. Proteiinia koodaavien osien lisäksi osa koodaavista alueista voi toimia säätelysekvensseinä pre-mRNA:ssa eksonisena silmukoinnin tehostajina tai eksonisen silmukoinnin silmukoinninvaimentimina.
Ei-trnasloituvat UTR-alueet
Transloimattomat alueet (UTR:t) ovat osia mRNA:sta ennen aloituskodonia ja lopetuskodonin jälkeisiä osia, joita ei transloida, ja niitä kutsutaan vastaavasti ”viisi prime transloimatonta aluetta” (5′ UTR) ja ”kolme prime ei-translatoitua aluetta” (3′ UTR).
Nämä alueet transkriptoidaan koodaavan alueen kanssa ja ovat siten eksonisia, koska ne ovat läsnä kypsässä mRNA:ssa. Geenin ilmentymisessä (geenin muuttamisessa proteiiniksi) useita tapahtuu transloimattomilla alueilla, mukaan lukien mRNA:n stabiilius, mRNA:n lokalisaatio ja translaatiotehokkuus.
UTR:n kyky suorittaa näitä toimintoja riippuu UTR:n sekvenssistä ja voi vaihdella mRNA:iden välillä. 3′ UTR:n geneettiset variantit on myös liitetty tautialttiuteen RNA-rakenteen ja proteiinien translaation muutoksen vuoksi [17]. mRNA:iden stabiilisuutta voidaan säädellä 5′-UTR:lla ja/tai 3′-UTR:lla, mikä johtuu vaihtelevasta affiniteetista RNA:ta hajottaville ribonukleaasi-entsyymeille ja apuproteiineille, jotka voivat edistää tai estää RNA:n hajoamista.
Translaatiotehokkuutta ja joskus täydellinen translaation esto, voidaan kontrolloida UTR:illä. Proteiinit, jotka sitoutuvat joko 3′- tai 5′-UTR:ään, voivat vaikuttaa translaatioon vaikuttamalla ribosomin kykyyn sitoutua mRNA:han. 3′ UTR:ään sitoutuneet mikroRNA:t voivat myös vaikuttaa translaatiotehokkuuteen tai mRNA:n stabiilisuuteen.
mRNA:n sytoplasmisen lokalisoinnin uskotaan olevan 3′-UTR:n funktio. Proteiinit, joita tarvitaan solun tietyllä alueella, voidaan myös kääntää siellä; sellaisessa tapauksessa 3′-UTR voi sisältää sekvenssejä, jotka sallivat transkriptin paikantamisen tälle alueelle translaatiota varten. Jotkut transloimattomien alueiden sisältämistä elementeistä muodostavat tunnusomaisen sekundaarirakenteen, kun ne transkriptoidaan RNA:ksi. Nämä rakenteelliset mRNA-elementit osallistuvat mRNA:n säätelyyn. Jotkut, kuten SECIS-elementti, ovat proteiinien sitoutumisen kohteita. Yksi mRNA-elementtiluokka, riboswitches, sitoo suoraan pieniä molekyylejä, muuttaen niiden laskostusta transkription tai translaation tasojen muuttamiseksi. Näissä tapauksissa mRNA säätelee itseään.
Poly(A) häntä
3′-poly(A)-häntä on pitkä sekvenssi adeniininukleotideja (usein useita satoja), jotka on lisätty pre-mRNA:n 3′-päähän. Tämä häntä edistää vientiä tumasta ja translaatiota sekä suojaa mRNA:ta hajoamiselta.
Monokistroninen ja polykistroninen mRNA
mRNA-molekyylin sanotaan olevan monokistroninen, kun se sisältää geneettisen tiedon vain yhden proteiiniketjun (polypeptidin) kääntämiseksi. Tämä koskee useimpia eukaryoottisia mRNA:ita [18][19]. Toisaalta polykistronisessa mRNA:ssa on useita avoimia lukukehyksiä (ORF), joista jokainen transloituu polypeptidiksi. Näillä polypeptideillä on yleensä läheinen toiminto (ne ovat usein alayksiköitä, jotka muodostavat lopullisen kompleksisen proteiinin) ja niiden koodaussekvenssi on ryhmitelty ja säädelty yhdessä säätelyalueella, joka sisältää promoottorin ja operaattorin. Suurin osa bakteereista ja arkeista löytyvästä mRNA:sta on monikistronista [18], kuten myös ihmisen mitokondriogenomi [20]. Dikistroninen tai bikistroninen mRNA koodaa vain kahta proteiinia.
mRNA:n sirkularisaatio (”ympyröiminen”)
Eukaryooteissa mRNA-molekyylit muodostavat pyöreitä rakenteita johtuen vuorovaikutuksesta eIF4E:n ja poly(A)-sitoutuvan proteiinin välillä, jotka molemmat sitoutuvat eIF4G:hen muodostaen mRNA-proteiini-mRNA-sillan [21]. Sirkularisaation (circularization) uskotaan edistävän ribosomien kiertokulkua mRNA:ssa, mikä johtaa aikatehokkaaseen translaatioon, ja se voi myös toimia varmistaen, että vain ehjä mRNA transloituu (osittain hajotulla mRNA:lla ei ole tyypillisesti m7G-huppua tai poly-A häntää) [22].
Muitakin sirkularisaatiomekanismeja on olemassa, erityisesti virusten mRNA:ssa. Polioviruksen mRNA käyttää apilanlehtiosaa 5′-päätään kohti PCBP2:n sitomiseen, joka sitoo poly(A)-sitovaa proteiinia muodostaen tutun mRNA-proteiini-mRNA-kehän. Ohran keltainen kääpiövirus sitoutuu mRNA-segmenttien väliin sen 5′- ja 3′-päässä (kutsutaan kissing stem-silmukoiksi), mikä kiertää mRNA:ta ilman minkäänlaisia proteiineja.
RNA-virusgenomit (joiden +-säikeet käännetään mRNA:ksi) ovat myös yleisesti ympyrämuotoisia. Genomin replikaation aikana sirkularisaatio tehostaa genomin replikaationopeuksia, ja oletetaan, että viruksen RNA-riippuvainen RNA-polymeraasi kiertää samalla tavalla kuin ribosomi.
mRNA:n hajoaminen
Saman solun eri mRNA:illa on erilainen elinkaari (stabiilisuus). Bakteerisoluissa yksittäiset mRNA:t voivat selviytyä sekunneista yli tuntiin. Niiden elinkaari on kuitenkin keskimäärin 1-3 minuuttia, mikä tekee bakteeri-mRNA:sta paljon epästabiilimman kuin eukaryoottinen mRNA[23].
Nisäkässoluissa mRNA:n elinkaari vaihtelee useista minuuteista päiviin [24]. Mitä suurempi mRNA:n stabiilius on, sitä enemmän proteiinia voidaan tuottaa mRNA:sta.
mRNA:n rajoitettu elinikä mahdollistaa solun muuttamaan proteiinisynteesiä nopeasti vastauksena muuttuviin tarpeisiinsa. On monia mekanismeja, jotka johtavat mRNA:n hajoamiseen.
Prokaryoottien mRNA: hajoaminen
Yleensä prokaryooteissa mRNA:n elinikä on paljon lyhyempi kuin eukaryooteissa. Prokaryootit hajottavat viestejä käyttämällä ribonukleaasien yhdistelmää, mukaan lukien endonukleaasit, 3′-eksonukleaasit ja 5′-eksonukleaasit.
Joissakin tapauksissa pienet RNA-molekyylit (sRNA), joiden pituus on kymmeniä – satoja nukleotideja, voivat stimuloida spesifisten mRNA:iden hajoamista muodostamalla emäsparin komplementaaristen sekvenssien kanssa ja helpottamalla ribonukleaasin pilkkoutumista RNaasi III:lla.
Äskettäin osoitettiin, että bakteereilla on myös eräänlainen 5′-huppu, joka koostuu trifosfaatista 5′-päässä [25]. Kahden fosfaatin poistaminen jättää 5′-monofosfaatin, mikä aiheuttaa mRNA:n hajoamiseen eksonukleaasi RNaasi J:n toimesta, joka hajoaa 5′:sta 3′:ksi.
Aitotumallisten mRNA-vaihtuvuus
Eukaryoottisoluissa vallitsee tasapaino translaatio- ja mRNA-hajoamisprosessien välillä. Aktiivisesti transloituvat viestit sitovat ribosomit, eukaryoottiset aloitustekijät eIF-4E ja eIF-4G sekä poly(A)-sitoutuva proteiini. eIF-4E ja eIF-4G estävät decapping-entsyymin (DCP2) ja poly(A)-sitoutuva proteiini estää eksosomikompleksin ja suojaa viestin päitä.
Translaation ja hajoamisen välinen tasapaino heijastuu P-kappaleina tunnettujen sytoplasmisten rakenteiden koossa ja runsaudessa [26]. mRNA:n poly(A) häntää lyhentävät erikoistuneet eksonukleaasit, jotka kohdistetaan tiettyihin lähetti-RNA:ihin cis-yhdistelmällä. – RNA:n säätelysekvenssit ja trans-vaikuttavat RNA:ta sitovat proteiinit. Poly(A) hännän poistamisen uskotaan häiritsevän viestin pyöreää rakennetta ja horjuttavan hupun sitomiskompleksia.
mRNA on sitten alttiina joko eksosomikompleksin tai hupunpoistokompleksin vaikutukselle. Tällä tavalla translaation kannalta epäaktiiviset mRNA:t voidaan tuhota nopeasti, kun taas aktiiviset mRNA:t pysyvät ennallaan. Mekanismia, jolla translaatio pysähtyy ja mRNA välitetään hajoamiskomplekseille, ei ymmärretä yksityiskohtaisesti.
AU-rikkaiden elementtien hajoaminen
AU:ta sisältävien elementtien läsnäolo joissakin nisäkkään mRNA:issa pyrkii epävakauttamaan näitä transkriptejä soluproteiinien vaikutuksesta, jotka sitovat näitä sekvenssejä ja stimuloivat poly(A) hännän poistoa.
Poly(A)-hännän häviämisen uskotaan edistävän mRNA:n hajoamista helpottamalla sekä eksosomikompleksin [27] että huppua poistavan (decapping) kompleksin hyökkäystä [28].
Nopea mRNA:n hajoaminen AU-rikkaiden elementtien kautta on kriittinen mekanismi voimakkaiden sytokiinien, kuten tuumorinekroositekijän (TNF) ja granulosyytti-makrofaagipesäkkeitä stimuloivan tekijän (GM-CSF) ylituotannon estämisessä [29].
AU-rikkaat elementit säätelevät myös protoonkogeenisten transkriptiotekijöiden, kuten c-Junin ja c-Fosin, biosynteesiä [30].
Nonsense-välitteinen rappeutuminen
Eukaryoottiset mRNA:t ovat nonsense-välitteisen rappeutumisen (NMD) -valvonnan alaisia. Se tarkistaa ennenaikaisten lopetuskodonien (nonsense-kodonien) läsnäolon mRNA:ssa. Nämä voivat johtua epätäydellisestä silmukoitumisesta, V(D)J-rekombinaatiosta adaptiivisessa immuunijärjestelmässä, mutaatioista DNA:ssa, transkriptiovirheistä, ribosomin vuotavasta skannauksesta, joka aiheuttaa kehyssiirtymän, ja muista syistä.
Ennenaikaisen lopetuskodonin havaitseminen laukaisee mRNA:n hajoamisen 5′-pään poistamisella, 3′-poly(A)-häntäpoistolla tai endonukleolyyttisellä katkaisulla [31].
Pienet häiritsevät RNA:t (siRNA)
Monisoluisten pienet häiritsevät RNA:t (siRNA:t) liitetään kompleksiin, joka tunnetaan nimellä RNA-indusoitu vaimennuskompleksi tai RISC. Tämä kompleksi sisältää endonukleaasin, joka pilkkoo täydellisesti komplementaarisia mRNA-molekyylejä, joihin siRNA sitoutuu. Tuloksena saadut mRNA-fragmentit tuhoavat sitten eksonukleaasit. siRNA:ta käytetään yleisesti laboratorioissa estämään geenien toimintaa soluviljelmässä. Sen uskotaan olevan osa luontaista immuunijärjestelmää suojana kaksijuosteisia RNA-viruksia vastaan [32].
MikroRNA (miRNA)
MikroRNA:t (miRNA:t) ovat pieniä RNA:ita, jotka tyypillisesti ovat osittain komplementaarisia monisoluisten lähetti-RNA:iden sekvensseille [33][34]. MiRNA:n sitoutuminen mRNA-molekyyliin voi tukahduttaa tämän mRNA:n translaation ja nopeuttaa poly(A) hännän poistoa, mikä nopeuttaa mRNA:n hajoamista. MiRNA:iden vaikutusmekanismi on aktiivisen tutkimuksen kohteena [35][36].
Muut rappeutumismekanismit
On olemassa muitakin tapoja, joilla mRNA-molekyylit voidaan hajottaa, mukaan lukien muun muassa non-stop hajoaminen ja hiljentäminen Piwin kanssa vuorovaikuttavan RNA:n (piRNA) avulla.
Sovellukset
RNA vaccine and RNA therapeutics
Nukleosidimodifioidun lähetti-RNA-sekvenssin antaminen voi saada solun valmistamaan proteiinia, joka puolestaan voisi suoraan hoitaa sairautta tai toimia rokotteena; epäsuoremmin proteiini voisi saada endogeenisen kantasolun erilaistumaan halutulla tavalla [37][38].
RNA-terapian ensisijaiset haasteet keskittyvät RNA:n toimittamiseen sopiviin soluihin [39]. Haasteisiin kuuluu se, että paljaat RNA-sekvenssit hajoavat luonnollisesti valmistuksen jälkeen; ne voivat laukaista kehon immuunijärjestelmän hyökkäämään niitä vastaan; ja ne eivät läpäise solukalvoa [38]. Kun ne ovat solun sisällä, niiden on poistuttava solun kuljetusmekanismista toimiakseen sytoplasmassa, jossa on tarvittavat ribosomit [37]. Käytössä olevissa koronarokotteissa kuljetus kohdesoluun ratkaistiin käyttämällä lipidipisaraa ja adenovirusvektoria (kuljetinta).
Näiden haasteiden voittamiseksi mRNA:ta terapeuttisena aineena esitettiin ensimmäisen kerran vuonna 1989 ”laajaisesti sovellettavan in vitro -transfektiotekniikan kehittämisen jälkeen”[40].
1990-luvulla kehitettiin eräitä syöpiä varten mRNA-rokotteita, jotka perustuvat ei-nukleosidimodifioituihin aineisiin. mRNA-pohjaisia hoitoja tutkitaan edelleen sekä syövän että autoimmuuni-, aineenvaihdunta- ja hengitystietulehdussairauksien hoitomenetelminä.
Geeniterapiat, kuten CRISPR, voivat myös hyötyä mRNA:n käytöstä indusoimaan soluja tuottamaan haluttua Cas-proteiinia [41]. Vuodesta 2010 lähtien RNA-rokotteita ja muita RNA-terapeuttisia aineita on kutsuttu ”seuraavan sukupolven lääkkeiksi”[42].
Ensimmäiset mRNA-pohjaiset rokotteet saivat rajoitetun luvan, ja Pfizer otti ne käyttöön kaikkialla maailmassa COVID-19-pandemian aikana. MRNA-teknologiaan perustuvat esimerkiksi nyt käytössä olevat BioNTech COVID-19 -rokote ja Moderna[43][44].
Viiteluettelo
- Cobb M (29 June 2015). ”Who discovered messenger RNA?”. Current Biology. 25 (13): R526–R532. doi:10.1016/j.cub.2015.05.032. PMID 26126273.
- Watson JD (February 22, 2013). Molecular Biology of the Gene, 7th edition. Pearson Higher Ed USA. ISBN 9780321851499.
- Choi YS, Patena W, Leavitt AD, McManus MT (March 2012). ”Widespread RNA 3′-end oligouridylation in mammals”. RNA (New York, N.Y.). 18 (3): 394–401. doi:10.1261/rna.029306.111. PMC 3285928. PMID 22291204.
- Quaresma AJ, Sievert R, Nickerson JA (April 2013). ”Regulation of mRNA export by the PI3 kinase/AKT signal transduction pathway”. Molecular Biology of the Cell. 24 (8): 1208–1221. doi:10.1091/mbc.E12-06-0450. PMC 3623641. PMID 23427269.
- Kierzkowski D, Kmieciak M, Piontek P, Wojtaszek P, Szweykowska-Kulinska Z, Jarmolowski A (September 2009). ”The Arabidopsis CBP20 targets the cap-binding complex to the nucleus, and is stabilized by CBP80”. The Plant Journal. 59 (5): 814–825. doi:10.1111/j.1365-313X.2009.03915.x. PMID 19453442.
- Strässer K, Masuda S, Mason P, Pfannstiel J, Oppizzi M, Rodriguez-Navarro S, Rondón AG, Aguilera A, Struhl K, Reed R, Hurt E (May 2002). ”TREX is a conserved complex coupling transcription with messenger RNA export”. Nature. 417 (6886): 304–308. Bibcode:2002Natur.417..304S. doi:10.1038/nature746. PMID 11979277. S2CID 1112194.
- Katahira J, Yoneda Y (27 October 2014). ”Roles of the TREX complex in nuclear export of mRNA”. RNA Biology. 6 (2): 149–152. doi:10.4161/rna.6.2.8046. PMID 19229134.
- Cenik C, Chua HN, Zhang H, Tarnawsky SP, Akef A, Derti A, Tasan M, Moore MJ, Palazzo AF, Roth FP (April 2011). ”Genome analysis reveals interplay between 5’UTR introns and nuclear mRNA export for secretory and mitochondrial genes”. PLOS Genetics. 7 (4): e1001366. doi:10.1371/journal.pgen.1001366. PMC 3077370. PMID 21533221.
- Steward O, Levy WB (March 1982). ”Preferential localization of polyribosomes under the base of dendritic spines in granule cells of the dentate gyrus”. The Journal of Neuroscience. 2 (3): 284–291. doi:10.1523/JNEUROSCI.02-03-00284.1982. PMC 6564334. PMID 7062109.
- Steward O, Worley PF (April 2001). ”Selective targeting of newly synthesized Arc mRNA to active synapses requires NMDA receptor activation”. Neuron. 30 (1): 227–240. doi:10.1016/s0896-6273(01)00275-6. PMID 11343657. S2CID 13395819.
- Job C, Eberwine J (December 2001). ”Localization and translation of mRNA in dendrites and axons”. Nature Reviews. Neuroscience. 2 (12): 889–898. doi:10.1038/35104069. PMID 11733796. S2CID 5275219.
- Hüttelmaier S, Zenklusen D, Lederer M, Dictenberg J, Lorenz M, Meng X, et al. (November 2005). ”Spatial regulation of beta-actin translation by Src-dependent phosphorylation of ZBP1”. Nature. 438 (7067): 512–5. doi:10.1038/nature04115. PMID 16306994. S2CID 2453397.
- Ainger K, Avossa D, Diana AS, Barry C, Barbarese E, Carson JH (September 1997). ”Transport and localization elements in myelin basic protein mRNA”. The Journal of Cell Biology. 138 (5): 1077–1087. doi:10.1083/jcb.138.5.1077. PMC 2136761. PMID 9281585.
- Lin CY, Beattie A, Baradaran B, Dray E, Duijf PH (September 2018). ”Contradictory mRNA and protein misexpression of EEF1A1 in ductal breast carcinoma due to cell cycle regulation and cellular stress”. Scientific Reports. 8 (1): 13904. Bibcode:2018NatSR…813904L. doi:10.1038/s41598-018-32272-x. PMC 6141510. PMID 30224719.
- Shabalina SA, Ogurtsov AY, Spiridonov NA (2006). ”A periodic pattern of mRNA secondary structure created by the genetic code”. Nucleic Acids Research. 34 (8): 2428–2437. doi:10.1093/nar/gkl287. PMC 1458515. PMID 16682450.
- Katz L, Burge CB (September 2003). ”Widespread selection for local RNA secondary structure in coding regions of bacterial genes”. Genome Research. 13 (9): 2042–2051. doi:10.1101/gr.1257503. PMC 403678. PMID 12952875.
- Lu YF, Mauger DM, Goldstein DB, Urban TJ, Weeks KM, Bradrick SS (November 2015). ”IFNL3 mRNA structure is remodeled by a functional non-coding polymorphism associated with hepatitis C virus clearance”. Scientific Reports. 5: 16037. Bibcode:2015NatSR…516037L. doi:10.1038/srep16037. PMC 4631997. PMID 26531896.
- Kozak M (March 1983). ”Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles”. Microbiological Reviews. 47 (1): 1–45. doi:10.1128/MMBR.47.1.1-45.1983. PMC 281560. PMID 6343825.
- Niehrs C, Pollet N (December 1999). ”Synexpression groups in eukaryotes”. Nature. 402 (6761): 483–487. Bibcode:1999Natur.402..483N. doi:10.1038/990025. PMID 10591207. S2CID 4349134.
- Mercer TR, Neph S, Dinger ME, Crawford J, Smith MA, Shearwood AM, Haugen E, Bracken CP, Rackham O, Stamatoyannopoulos JA, Filipovska A, Mattick JS (August 2011). ”The human mitochondrial transcriptome”. Cell. 146 (4): 645–658. doi:10.1016/j.cell.2011.06.051. PMC 3160626. PMID 21854988.
- Wells SE, Hillner PE, Vale RD, Sachs AB (July 1998). ”Circularization of mRNA by eukaryotic translation initiation factors”. Molecular Cell. 2 (1): 135–140. CiteSeerX 10.1.1.320.5704. doi:10.1016/S1097-2765(00)80122-7. PMID 9702200.
- López-Lastra M, Rivas A, Barría MI (2005). ”Protein synthesis in eukaryotes: the growing biological relevance of cap-independent translation initiation”. Biological Research. 38 (2–3): 121–146. doi:10.4067/S0716-97602005000200003. PMID 16238092.
- Lewin B, Krebs JE, Kilpatrick ST, Goldstein ES, eds. (2011). Lewin’s genes X (10th ed.). Sudbury, Mass.: Jones and Bartlett. ISBN 9780763766320. OCLC 456641931.
- Yu J, Russell JE (September 2001). ”Structural and functional analysis of an mRNP complex that mediates the high stability of human beta-globin mRNA”. Molecular and Cellular Biology. 21 (17): 5879–5888. doi:10.1128/mcb.21.17.5879-5888.2001. PMC 87307. PMID 11486027.
- Deana A, Celesnik H, Belasco JG (January 2008). ”The bacterial enzyme RppH triggers messenger RNA degradation by 5′ pyrophosphate removal”. Nature. 451 (7176): 355–358. Bibcode:2008Natur.451..355D. doi:10.1038/nature06475. PMID 18202662. S2CID 4321451.
- Parker R, Sheth U (March 2007). ”P bodies and the control of mRNA translation and degradation”. Molecular Cell. 25 (5): 635–646. doi:10.1016/j.molcel.2007.02.011. PMID 17349952.
- Chen CY, Gherzi R, Ong SE, Chan EL, Raijmakers R, Pruijn GJ, Stoecklin G, Moroni C, Mann M, Karin M (November 2001). ”AU binding proteins recruit the exosome to degrade ARE-containing mRNAs”. Cell. 107 (4): 451–464. doi:10.1016/S0092-8674(01)00578-5. PMID 11719186. S2CID 14817671.
- Fenger-Grøn M, Fillman C, Norrild B, Lykke-Andersen J (December 2005). ”Multiple processing body factors and the ARE binding protein TTP activate mRNA decapping” (PDF). Molecular Cell. 20 (6): 905–915. doi:10.1016/j.molcel.2005.10.031. PMID 16364915. Archived from the original (PDF) on 2011-06-06.
- Shaw G, Kamen R (August 1986). ”A conserved AU sequence from the 3′ untranslated region of GM-CSF mRNA mediates selective mRNA degradation”. Cell. 46 (5): 659–667. doi:10.1016/0092-8674(86)90341-7. PMID 3488815. S2CID 40332253.
- Chen CY, Shyu AB (November 1995). ”AU-rich elements: characterization and importance in mRNA degradation”. Trends in Biochemical Sciences. 20 (11): 465–470. doi:10.1016/S0968-0004(00)89102-1. PMID 8578590.
- Isken O, Maquat LE (August 2007). ”Quality control of eukaryotic mRNA: safeguarding cells from abnormal mRNA function”. Genes & Development. 21 (15): 1833–1856. doi:10.1101/gad.1566807. PMID 17671086.
- Obbard DJ, Gordon KH, Buck AH, Jiggins FM (January 2009). ”The evolution of RNAi as a defence against viruses and transposable elements”. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1513): 99–115. doi:10.1098/rstb.2008.0168. PMC 2592633. PMID 18926973.
- Robert E. Farrell, Jr. RNA Methodologies, 5th Edition. Academic Press, 2017
- Brennecke J, Stark A, Russell RB, Cohen SM (March 2005). ”Principles of microRNA-target recognition”. PLOS Biology. 3 (3): e85. doi:10.1371/journal.pbio.0030085. PMC 1043860. PMID 15723116.
- Tasuku Honjo, Michael Reth, Andreas Radbruch, Frederick Alt. Molecular Biology of B Cells, 2nd Edition. Academic Press, 2014 (including ”updated research on microRNAs”)
- Eulalio A, Huntzinger E, Nishihara T, Rehwinkel J, Fauser M, Izaurralde E (January 2009). ”Deadenylation is a widespread effect of miRNA regulation”. RNA. 15 (1): 21–32. doi:10.1261/rna.1399509. PMC 2612776. PMID 19029310.
- Hajj KA, Whitehead KA (12 September 2017). ”Tools for translation: non-viral materials for therapeutic mRNA delivery”. Nature Reviews Materials. 2 (10): 17056. Bibcode:2017NatRM…217056H. doi:10.1038/natrevmats.2017.56.
- Gousseinov E, Kozlov M, Scanlan C (September 15, 2015). ”RNA-Based Therapeutics and Vaccines”. Genetic Engineering News.
- Kaczmarek JC, Kowalski PS, Anderson DG (June 2017). ”Advances in the delivery of RNA therapeutics: from concept to clinical reality”. Genome Medicine. 9 (1): 60. doi:10.1186/s13073-017-0450-0. PMC 5485616. PMID 28655327.
- Schlake T, Thess A, Fotin-Mleczek M, Kallen KJ (November 2012). ”Developing mRNA-vaccine technologies”. RNA Biology. 9 (11): 1319–30. doi:10.4161/rna.22269. PMC 3597572. PMID 23064118.
- Haridi R (2021-04-23). ”The mRNA revolution: How COVID-19 hit fast-forward on an experimental technology”. New Atlas. Retrieved 2021-04-26.
- Kowalska J, Wypijewska del Nogal A, Darzynkiewicz ZM, Buck J, Nicola C, Kuhn AN, Lukaszewicz M, Zuberek J, Strenkowska M, Ziemniak M, Maciejczyk M, Bojarska E, Rhoads RE, Darzynkiewicz E, Sahin U, Jemielity J (2014), ”mRNA-based therapeutics–developing a new class of drugs.”, Nature Reviews Drug Discovery, 13 (10), pp. 759–780, doi:10.1093/nar/gku757, PMC 4176373, PMID 25150148
- P. Polack, Fernando; Thomas, Stephen J.; Kitchin, Nicholas; Absalon, Judith; Gurtman, Alejandra; Lockhart, Stephen; Perez, John L.; Pérez Marc, Gonzalo; Moreira, Edson D.; Zerbini, Cristiano; Bailey, Ruth; Swanson, Kena A.; Roychoudhury, Satrajit; Koury, Kenneth; Li, Ping; Kalina, Warren V.; Cooper, David; Frenck, Robert W.; Hammitt, Laura L.; Türeci, Özlem; Nell, Haylene; Schaefer, Axel; Ünal, Serhat; Tresnan, Dina B.; Mather, Susan; Dormitzer, Philip R.; Şahin, Uğur; Jansen, Kathrin U.; Gruber, William C. (2020), ”Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine.”, New England Journal of Medicine (383): 2603–15, doi:10.1056/NEJMoa2034577
- Walsh, Edward E.; Frenck, Robert W.; Falsey, Ann R.; Kitchin, Nicholas; Absalon, Judith; Gurtman, Alejandra; Lockhart, Stephen; Neuzil, Kathleen; Mulligan, Mark J.; Bailey, Ruth; Swanson, Kena A.; Li, Ping; Koury, Kenneth; Kalina, Warren; Cooper, David; Fontes Garfias, Camila; Shi, Pei Yong; Türeci, Özlem; Tompkins, Kristin R.; Lyke, Kirsten E.; Raabe, Vanessa; Dormitzer, Philip R.; Jansen, Kathrin U.; Şahin, Uğur; Gruber, William C. (2020), ”Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates.”, New England Journal of Medicine (383): 2439–50, doi:10.1056/NEJMMoa2027906
Aiheesta enemmän
- Alsaweed M, Lai CT, Hartmann PE, Geddes DT, Kakulas F (February 2016). ”Human milk miRNAs primarily originate from the mammary gland resulting in unique miRNA profiles of fractionated milk”. Scientific Reports. 6 (1): 20680. Bibcode:2016NatSR…620680A. doi:10.1038/srep20680. PMC 4745068. PMID 26854194.
- Lillycrop KA, Burdge GC (October 2012). ”Epigenetic mechanisms linking early nutrition to long term health”. Best Practice & Research. Clinical Endocrinology & Metabolism. 26 (5): 667–676. doi:10.1016/j.beem.2012.03.009. PMID 22980048.
- Melnik BC, Kakulas F, Geddes DT, Hartmann PE, John SM, Carrera-Bastos P, Cordain L, Schmitz G (21 June 2016). ”Milk miRNAs: simple nutrients or systemic functional regulators?”. Nutrition & Metabolism. 13 (1): 42. doi:10.1186/s12986-016-0101-2. PMC 4915038. PMID 27330539.
- Vickers MH (June 2014). ”Early life nutrition, epigenetics and programming of later life disease”. Nutrients. 6 (6): 2165–2178. doi:10.3390/nu6062165. PMC 4073141. PMID 24892374.
- Zhou Q, Li M, Wang X, Li Q, Wang T, Zhu Q, Zhou X, Wang X, Gao X, Li X (2012). ”Immune-related microRNAs are abundant in breast milk exosomes”. International Journal of Biological Sciences. 8 (1): 118–123. doi:10.7150/ijbs.8.118. PMC 3248653. PMID 22211110.
Lähde: https://en.wikipedia.org/wiki/Messenger_RNA