Insuliini ja terveys: Kolmas luku
Kuvasin kahdessa edellisessä artikkelissa (Insuliini ja terveys: Johdanto & Insuliini ja terveys: Hiilihydraatti-insuliinimalli) insuliinin toimintaperiaatteita ja vaikutuksia yleisellä tasolla. Insuliini on elintärkeä hormoni, joka vaikuttaa moniin aineenvaihduntatapahtumiin.
Insuliiniresistenssi liittyy useimpiin elintapasairauksiin lihavuudesta aikuistyypin diabetekseen. Amerikkalaisista jo lähes puolet sairastaa tietämättään insuliiniresistenssiä.
Insuliiniresistenssi ylläpitää nälkää ja lihomista kahdella tavalla:
- Veressä on syömisen jälkeen runsaasti glukoosia, mutta insuliiniresistenttien solujen glukoosinotto on häiriintynyt, jolloin ne eivät pysty hyödyntämään glukoosia energian tuotannossa. Veren sokeripitoisuus pysyy korkeana.
- Tämän vuoksi insuliini päästää verestä glukoosia rasvasoluihin, joiden insuliiniherkkyys säilyy pidempään kuin lihasten insuliiniherkkyys. Rasvasoluissa glukoosi muutetaan triglyserideiksi, eli läskiksi.
Insuliiniresistenssi ylläpitää nälkää, koska solut eivät pysty tuottamaan glukoosista riittävästi energiaa ja elimistö reagoi solujen energianpuutteeseen hormonaalisesti.
Verestä glukoosi on siirrettävä johonkin, koska korkea verensokeri kasvattaa glykaation riskiä. Glykaatiossa glukoosi sokeroi aminohappoja ja rasvahappoja, jolloin niistä tulee melkein siirappia. Se ei ole ollenkaan toivottavaa.
Glykaation tuottamat AGE:t (Advanced Glycation End-products), eli glykaation lopputuotteet kasvattavat sairastumisen riskiä.
Myös diabetekseen liittyvä virtsaamistarpeen lisääntyminen liittyy siihen, että elimistö yrittää päästä eroon ylimääräisestä sokerista. Diabetes tunnettiin aluksi makeavirtsaisuutena ja monet lääkärit varmistuivat diabetes-diagnoosista haistamalla tai maistamalla potilaansa virtsaa.
Insuliinia käytetään yhä yleisemmin doping-aineena
Yhä useammat urheilijat ja kuntoilijat käyttävät insuliinia suorituskyvyn parantamiseksi, kertoo New Scientist. Insuliinia on käytännössä melkein mahdotonta havaita verestä dopingtesteillä.
Väärin annosteltuna insuliini voi tappaa
Kehonrakentajat olivat edelläkävijöitä insuliinin doping-käytössä jo vuosia sitten. Näyttöä insuliinin laajemmasta hyväksikäytöstä ei ole aiemmin juuri saatu, mutta näyttö on viime aikoina lisääntynyt erityisesti kuntoilijoille ja kehonrakentajille tarkoitetun neulanvaihtojärjestelmän kokoamien tietojen avulla.
Insuliini auttaa urheilijoita kahdella tavalla:
Kehonrakentajilla insuliini toimii anabolisten steroidien, kuten testosteronin tai ihmisen kasvuhormonin rinnalla lihaskudoksen vahvistajana. Steroidit rakentavat lihaskudosta ja insuliini estää lihasproteiinien hajoamisen.
Insuliini vahvistaa myös mm. keskimatkan juoksijoiden ja muiden urheilijoiden kestävyyttä mahdollistamalla tehokkaamman lihasglykogeenien sokeritankkauksen.
Mitä suuremmat glykogeenivarastot lihaksilla on, sitä kauemmpin lihakset jaksavat puurtaa väsymättä. Kestävyysurheilijoilla tankkaus voi vaikuttaa kilpailun lopputulokseen.
Tankatessa urheilijoiden on infusoitava insuliinia ja glukoosia samanaikaisesti muutaman tunnin ajan vereen. Menetelmä aiheuttaa tarkoituksellisen hyperinsulinemian.
Pitkällä aikavälillä anabolisten steroidien käyttö voi vahingoittaa mm. lisääntymisterveyttä ja mieskuntoa. Insuliinin yliannostus voi myös aiheuttaa kuolemaan johtavan kooman poistamalla verestä niin paljon sokeria, että aivosolut eivät saa riittävästi energiaa ja happea.
Houkutus insuliinin käyttöön on kuitenkin suuri. Sen teho on ilmeinen ja se häviää nopeasti verenkierrosta. Noin puolet insuliinista poistuu verenkierrosta vain neljässä minuutissa. Vaikka insuliinia havaittaisiin, sitä ei voida mitenkään erottaa ihmisen omasta insuliinista.
Hyperinsulinemia voi kasvattaa glukoosimetabolian nopeutta kaksinkertaiseksi
Kansainvälinen olympiakomitea kielsi insuliinin vuonna 1998. Kielto ei kuitenkaan koske diabeetikkoja, joiden terveys riippuu insuliinista. Diabetes UK, joka edustaa maan diabeetikkoja, myöntää, että joillakin diabeetikoilla saattaa olla taloudellinen houkutus myydä insuliinia doping-käyttöön.
Insuliini on anabolinen hormoni
Insuliini on anabolinen hormoni, jolla on metabolisia vaikutuksia koko kehossa. Haiman Langerhansin saarekkeiden beetasolut vastaavat insuliinisynteesistä.
Beetasolut säätelevät insuliinin tuotantoa seuraamalla plasman sisällä kiertävien energiaravinteiden, glukoosin, aminohappojen, ketohappojen ja rasvahappojen tasoja.
Insuliini orkestroi energiansäästöä ja -käyttöä ravinnonsaannin ja paaston aikana.
Insuliiniaineenvaihdunnan käsitteitä
- Glukoneogeneesi: Glukoosin syntetisoiminen eräistä aminohapoista, glyserolista ja sitruunahappokierron väliaineista
- Glykolyysi: Glukoosin hajoaminen pyruviinihapoksi* sekä energian tuottaminen glukoosista
- Glykogeneesi: Glykogeenin synteesi glukoosista
- Glykogenolyysi: Glykogeenin hajoaminen glukoosiksi
- Lipogeneesi: Asetyylikoentsyymi-A:n muuttaminen rasvahapoiksi ja sitä seuraava triglyseridisynteesi
- Lipolyysi: Lipidien ja triglyseridien hajoaminen vapaiksi rasvahapoiksi ja glyseroliksi. Vapaista rasvahappoista valmistetaan ketoaineita (asetoni, asetoasetaatti ja betahydroksibutyraatti). Glyserolia käytetään glukoneogeneesissä glukoosin syntetisoimiseen.
Pyruviinihappo ja pyruvaatti
*Termejä pyruvaaatti (pyruvate) ja pyruviinihappoa (pyruvic acid) käytetään usein rinnakkain. Niiden välillä on selvä ero. Pyruviinihappo on happo, joka voi vapauttaa vetyionin ja sitoutua positiivisesti varautuneeseen natrium- tai kaliumioniin happosuolan muodostamiseksi.
Happosuola tunnetaan nimellä pyruvaatti. Toisin sanoen pyruvaatti on pyruviinihapon suola tai esteri.
Keskeinen ero pyruviinihapon ja pyruvaatin välillä on, että karboksyylihapporyhmän vetyatomi puuttuu tai se on poistettu. Pyruviinihapon heikon happamuusluonteen vuoksi se dissosioituu helposti vedessä ja muodostaa siten pyruvaatin.
Pyruvaatti on tärkeä kemiallinen yhdiste ihmisen aineenvaihdunnassa ja biokemiassa. Pyruvaatti osallistuu glukoosin metaboliaan, eli glykolyysiin, jossa yksi glukoosimolekyyli hajoaa kahdeksi pyruvaattimolekyyliksi. Pyruvaattimolekyylejä käytetään edelleen muissa reaktiosarjoissa (sitruunahappokierrossa) energian tuottamiseksi.
Insuliinin solumetabolia
Lihaskudoksen, verisuonten endoteelin, sydämen ja maksan soluissa tapahtuu insuliinin aktivoima kaskadi.
Biokemiallinen kaskadi tunnetaan myös signalointikaskadina tai signalointireitinä. Kaskadi tarkoittaa yleensä jonkin ärsykkeen laukaisemia soluissa lineaarisesti eteneviä kemiallisia reaktioita. Suurin osa biokemiallisista kaskadeista on tapahtumasarjoja, joissa yksi biokemiallinen tapahtuma laukaisee seuraavan jne.
Insuliinin tuottama vaste näissä soluissa on kudosspesifinen. Rasvakudoksessa, luurankolihaksissa ja sydämessä insuliinin aktivoima kaskadi johtaa glukoosimetaboliaan,josa solut ottavat vastaan glukoosia.
Vasodilataatio ja erektion helppous
Vasodilataatio, eli verisuonten laajeneminen typpioksidin (NO) tuotannon kautta on insuliinisignaloinnin seuraus verisuonten endoteelissa ja sydämessä.
Typpioksidi ja vasodilataatio näyttelevät tärkeää roolia myös makuuhuoneissa vaikuttamalla erektioon.
Insuliiniresistenssi ennakoi aikuistyypin diabetesta, joka tunnetusti aiheuttaa erektiohäiriöitä, mutta insuliiniresistenssi on myös täysin itsenäinen erektiohäiriöiden aiheuttaja jo ennen diabetekseen sairastumista.
Erektiohäiriöt lisääntyvät selvästi 40 ikävuoden jälkeen. Erektiohäiriöiden syy ei ole itsenäinen sairaus, vaan siihen vaikuttavat erityisesti:
- verenkiertoelinten sairaudet ja niiden hoitoon käytetyt lääkkeet
- insuliiniresistenssi
- metabolinen oireyhtymä
- diabetes
- lihavuus
- tupakointi
- testosteronipitoisuuden lasku
- kilpirauhasen ali- tai ylitoiminta
- neurologiset sairaudet
PDE5-estäjät, kuten sildenafiili, tadalafiili, vardenafiili ja avanafiili parantavat erektiota lisäämällä solunsisäistä syksista guanosiinimonofosfaattia, jolloin peniksen paisuvaiskudoksen, virtsarakon, eturauhasen ja virtsaputken sileä lihas rentoutuu.
Lyhyestä virsi kaunis: Seksi on kivaa. Lihominen, insuliiniresistenssi, metabolinen oireyhtymä ja diabetes tekevät seksistä kuitenkin hankalaa tai mahdotonta. Näiden ongelmien esiintyminen on signaali, johon miehen tulee kiinnittää huomiota. Erektiovaikeus voi kertoa sydän- ja verisuonitaudeista, diabeteksesta tai jostakin muusta sairaudesta.
Tämä vaiettu ongelma on hyvin yleinen. Noin 200 000 suomalaismiestä kärsii vaikeasta tai keskivaikeasta erektiohäiriöstä. 300 000 suomalaismiehellä on lievä erektiohäiriö. Pieni elintapojen korjaus voi siis parantaa terveyttä monin tavoin.
Kerrataan: Insuliiniresistenssi ja hyperinsulinemia ovat sydän- ja verisuonitautien itsenäisiä riskitekijöitä. Fokusointi LDL-kolesterolin vähentämiseen on johtanut tilanteeseen, jossa ihmiset sairastuvat ja kuolevat kardiometabolisiin sairauksiin, jotka saataisiin lääkkeettömään remissioon verensokeria ja insuliinia laskevilla pienillä elintapojen korjauksilla.
Aineenvaihdunta
Maksassa insuliinin määrä vaikuttaa glukoneogeneesiin, joka hidastuu ja loppuu. Sen sijaan glukoosin varastoiminen glykogeeneihin glykogeneesissä lisääntyy insuliinin vaikutuksesta.
Insuliinin vaikutus ulottuu myös lipidi- ja proteiinimetaboliaan. Se stimuloi lipogeneesiä ja proteiinisynteesiä ja päinvastoin estää lipolyysiä ja proteiinin hajoamista.
Molekyylinebiologia: tekninen ja huonosti suomennettu osa!
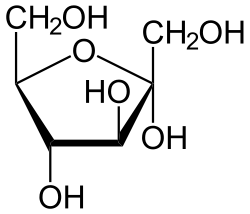
Insuliini on peptidihormoni, joka koostuu 51 aminohaposta, jotka jakautuvat kahden peptidiketjun, 21 ja 30 aminohappotähteen A- ja B-ketjuiksi.
Kysteiinitähteiden disulfidisidokset yhdistävät nämä kaksi ketjua. Preproinsuliini on insuliinin alkuperäinen esiasteproteiini. Se on yksiketjuinen polypeptidi, joka koostuu proinsuliinista ja signaalipeptidisekvenssistä. Sen siirtyessä endoplasmiseen retikulumiin preproinsuliini pilkkoutuu sen signaalipeptidistä vapauttaen proinsuliinia. Yksiketjuinen proinsuliini sisältää A- ja B-ketjut ja C-peptidin (tai C-domeenin), kuten kuvassa.
 C-peptidi muodostuu haiman Langerhansin saarekkeiden β-soluissa syntetisoidusta proinsuliinista sen pilkkoutuessa insuliiniksi ja C-peptidiksi. Sitä erittyy vereen insuliinin kanssa ekvimolaarisina määrinä.
C-peptidi muodostuu haiman Langerhansin saarekkeiden β-soluissa syntetisoidusta proinsuliinista sen pilkkoutuessa insuliiniksi ja C-peptidiksi. Sitä erittyy vereen insuliinin kanssa ekvimolaarisina määrinä.
Kaksiemäksiset tähteet reunustavat C-domeenia kummassakin päässä. Jokaisen kaksiemäksisen jäännöksen kohdalla trypsiinin kaltainen entsyymi katkaisee proinsuliinin.
Tämä vapauttaa lopulta insuliinin ja C-peptidin. Insuliinia varastoidaan sinkki-insuliiniheksameereinä glukoosille herkissä eritysrakkuloissa, kunnes sitä tarvitaan metabolisesti.
Muuttumattoman proinsulinin määrä korreloi merkitsevästi insuliiniresistenssin kanssa. Adiponektiiniarvot ovat insuliiniresistenteillä hieman normaalia alhaisemmat, mutta resistiini ei näytä korreloivan insuliiniresistenssin kanssa. Kohonnut proinsuliini kuvastaa pitkälle edennyttä solujen vauriota ja on hyvin spesifinen insuliiniresistenssin indikaattori. – Terve.fi
Insuliinin merkitys glukoosimetaboliassa
Glukoosimetabolian homeostaasia ylläpidetään kahdella signalointikaskadilla. Ne ovat: insuliinin välittämä glukoosinotto (IMGU) ja glukoosistimuloitu insuliinin eritys (GSIS).
IMGU-kaskadi antaa insuliinin lisätä glukoosin imeytymistä luurankolihaksissa ja rasvakudoksessa sekä estää glukoosin muodostumista maksasoluissa.
Insuliinikaskadin alavirran signaloinnin aktivointi alkaa, kun solun ulkopuolinen insuliini on vuorovaikutuksessa insuliinireseptorin alfa-alayksikön kanssa. Tämä vuorovaikutus johtaa konformaatiomuutoksiin insuliinireseptorikompleksissa, mikä edelleen johtaa insuliinireseptorisubstraattien tyrosiinikinaasifosforylaatioon ja sen jälkeiseen fosfatidyyli-inositoli-3-kinaasin aktivoitumiseen.
Fosfatidyyli-inositoli on fosfolipidi, joka koostuu glyserolista, rasvahaposta ja inositolista. Fosfatidyyli-inositoli ja sen fosforyloidut johdokset ovat solujen toisiolähettejä, jotka osallistuvat solun toiminnan säätelyyn. Fosfatidyyli-inositoli-4,5-bisfosfaatin plasmamebraanipää toimii solutukirangan proteiinien sekä joidenkin eksosytoosissa tarvittavien proteiinien kiinnityskohtana. Lipidi osallistuu myös solun signaalinvälitykseen. Tällöin se vapautuu solun reagoidessa solun ulkoiseen signaaliin tiettyjen reseptorien välityksellä. Signaalinvälitys tapahtuu reaktiosarjassa, joka alkaa lipidin fosfaattipään poistamisella ja päättyy proteiinikinaasi C:n aktivaatioon. – Wikipedia
Nämä alavirran tapahtumat aktivoivat GLUT-4-kuljettusmolekyylin siirtymisen solun plasmakalvolle. Solunsisäisesti GLUT4:ää esiintyy rakkuloissa. Näiden GLUT4-rakkuloiden eksosytoitumisnopeus kasvaa insuliinin toiminnan vuoksi. Lisäämällä GLUT-4-kuljetttimien läsnäoloa plasmakalvossa, insuliini sallii glukoosin pääsyn luurankolihassoluihin, jossa se voi metaboloitua glykolyysissä pyruvaateiksi tai varastoitua glykogeeniksi.
Insuliinin rooli glykogeenimetaboliassa
Maksassa insuliini vaikuttaa glykogeenimetaboliaan stimuloimalla glykogeenisynteesiä. Proteiinifosfataasi I (PPI) on avainmolekyyli glykogeenimetabolian säätelyssä.
Defosforylaation kautta PPI hidastaa glykogenolyysinopeutta inaktivoimalla fosforylaasikinaasin ja fosforylaasi A:n. Sitä vastoin PPI kiihdyttää glykogeneesiä aktivoimalla glykogeenisyntaasi B:n. Insuliini lisää PPI-substraattikohtaista aktiivisuutta glykogeenipartikkeleille puolestaan stimuloimalla glykogeenin synteesiä maksan glukoosista.
Insuliini hallitsee suoraan erilaisia maksan metabolisia entsyymejä geenitranskriptiolla. Tämä vaikuttaa geenien ilmentymiseen aineenvaihduntareiteillä
Glukoneogeneesissä insuliini estää nopeutta rajoittavan fosfoenolipyruvaattikarboksylaasin, samoin kuin fruktoosi-1,6-bisfosfataasin ja glukoosi-6-fosfataasin geeniekspressiota.
Glykolyysissä glukokinaasin ja pyruvaattikinaasin geeniekspressio kasvaa. Lipogeneesissä ilmentyminen lisääntyy rasvahapposyntaasista, pyruvaattidehydrogenaasista ja asetyyli-CoA-karboksylaasista.
Insuliinin rooli lipidien aineenvaihdunnassa
Kuten aiemmin todettiin, insuliini lisää joidenkin lipogeenisten entsyymien ilmentymistä. Tämä johtuu rasvasoluihin eli adiposyytteihin lipidiksi varastoituneesta glukoosista. Rasvasolujen kasvu lisää adiposyyttien glukoosinottoa.
Insuliini säätelee tätä prosessia edelleen defosforyloimalla ja estämällä sen jälkeen hormoniherkän lipaasin (HSL) erittymistä, mikä johtaa lipolyysin estoon. Viime kädessä insuliini vähentää näin seerumin vapaiden rasvahappojen määrää.
Tällä on välitön vaikutus painonhallintaan. Insuliini estää rasvasoluihin varastoidun energian hyödyntämisen estämällä triglyseridejä vapaiksi rasvahapoiksi pilkkovan lipolyysin kannalta välttämättömän hormoniherkän lipaasin vaikutuksen.
Lipolyysi pilkkoo rasvasoluihin varastoituneita triglyseridejä vapaiksi rasvahapoiksi ja glyseroliksi. Vapaista rasvahapoista maksa tuottaa ketoaineita. Glyserolia käytetään glukoneogeneesissä, joka tuottaa glukoosia. Ajan mittaan solut oppivat tuottamaan vapaista rasvahapoista energiaa beetaoksidaatiossa.
Insuliinin rooli proteiinien aineenvaihdunnassa
Insuliini säätelee osittain proteiinien vaihtuvuutta. Lyhytketjuisten aminohappojen, kuten alaniinin, arginiinin ja glutamiinin soluunotto lisääntyy insuliinin vaikutuksesta. Tämä stimuloi proteiinisynteesiä.
Protein turnover rate is regulated in part by insulin. Protein synthesis is stimulated by insulin’s increase in intracellular uptake of alanine, arginine, and glutamine (short chain amino acids), as well as gene expression of albumin and muscle myosin heavy chain alpha.
Regulation of protein breakdown is affected by insulin’s downregulation of hepatic and muscle cell enzymes responsible for protein degradation. The impacted enzymes include ATP-ubiquitin-dependent proteases, and ATP-independent lysosomal proteases and hydrolases.
Insuliinin rooli tulehduksessa ja vasodilataatiossa
Insuliini vaikuttaa verisuonten endoteelisoluissa ja makrofageissa anti-inflammatorisesti, eli tulehdusta ehkäisevästi. Endoteelisoluissa insuliini stimuloi endoteelin typpioksidisyntaasin (eNOS) ilmentymistä. eNOS toimii vapauttamalla typpioksidia (NO), mikä johtaa verisuonten laajenemiseen.
Insuliini vaimentaa endoteelisolujen tumatekijä-kappa-B:tä (NF-kB). Endoteelin NF-KB aktivoi adheesiomolekyylien, E-selektiinin ja ICAM-1:n ilmentymisen, joka vapauttaa liukoisia soluadheesiomolekyylejä verenkiertoon.
Tutkimukset ovat yhdistäneet solujen adheesiomolekyylien esiintymisen verisuonten endoteelissa valtimoiden ateroskleroottisten plakkien muodostumiseen ja sydäntauteihin.
Insuliini estää reaktiivisten happiradikaalien (ROS) muodostumisen. Makrofagissa insuliini estää NADPH-oksidaasin ilmentymistä tukahduttamalla yhtä sen avainkomponenteista, p47phoxia.
NADPH-oksidaasia tarvitaan synnyttämään happiradikaaleja, jotka aktivoivat NF-kB:n estäjää, eli kinaasibeeta-inhibiittoria (IKKB). IKKB fosforyloi IkB:n, mikä johtaa sen hajoamiseen. Tämä hajoaminen vapauttaa NF-kB:n, mikä mahdollistaa sen translokaation makrofagin ytimessä.
Solun tumassa ollessaan NF-kB stimuloi verenkiertoon vapautuvien tulehdusta edistävien proteiinien geenien transkriptiota:
- indusoituva typpioksidisyntaasi (iNOS)
- tuumorinekroositekijä-alfa (TNF-alfa)
- interleukiini-6 (IL -6)
- interleukiini-8 (IL-8)
- monosyyttien kemoattraktantiproteiini (MCP-1)
- matriisimetalloproteinaasi (MMP)
Lääketieteellinen merkitys
Monet metaboliset sairaudet liittyvät krooniseen hyperglykemiaan, hyperinsulinemiaan ja insuliiniresistenssiin.
Tyypin 1 insuliinista riippuvainen diabetes mellitus (DM1) on tauti, jossa haiman insuliinin tuotanto on vähäistä tai kokonaan loppunut. Koska insuliinia tarvitaan solujen glukoosinottoon, diabeteksessa solujen kyky tuottaa glukoosista energiaa romahtaa. Tämän seurauksena hoitamaton tyypin 1 diabetes johtaa nälkiintymiseen ja kuolemaan.
Tyypin 2 insuliinista riippumaton diabetes mellitus (DM2) on metabolinen sairaus, jossa keho tuottaa yhä insuliinia, mutta jatkuvan hyperglykemian (korkean verensokerin) vuoksi solujen glukoosinotto vuosien mittaan heikentyy. Tämä tarjonnan ja kysynnän epäsuhta johtaa insuliiniresistenssiin ja epänormaaliin glukoosimetaboliaan.
Molemmat diabetekset ovat sokeriaineenvaihdunnan häiriöitä. Ruokailun jälkeen verensokeri pysyy koholla, koska haima ei tuota insuliinia (DM1) tai solujen insuliiniherkkyys on vaurioitunut (DM2). Kohollaan oleva verensokeri ja korkeat veren insuliinipitoisuudet vahingoittavat verisuonia ja elimiä. Tyypin 2 diabetes lisää erektiovaikeuksia, lihomista ja komplikaatioita, joihin kuuluvat mm. neuropatia, munuaisten vajaatoiminta, retinopatia, sydän- ja verisuonitaudit sekä perifeeriset verisuonisairaudet.
Tyypin 2 diabeteksen alkuvaiheessa haiman beetasolut tuottavat riittävästi insuliinia energia-aineenvaihdunnan ylläpitämiseksi, mutta insuliinin tuotanto muuttuu tehottomaksi, kun osa insuliinin välittämästä glukoosinottokaskadista ei enää toimi.
Erityisesti glukoosin kulkeutuminen plasmamembraanin läpi GLUT-4-kuljettimien kautta heikentyy, mikä kertoo solujen insuliiniresistenssista. Insuliiniresistenssi vaikuttaa ensin lihassoluihin. Rasvasolujen insuliinisensitiivisyys pysyy yleensä hyvänä varsin pitkään, minkä vuoksi glukoosia päätyy rasvasoluihin lihasten sijaan.
Pahoittelen, jos tekstiin jäi käännös- ja/tai asiavirheitä.

https://www.ncbi.nlm.nih.gov/books/NBK525983/