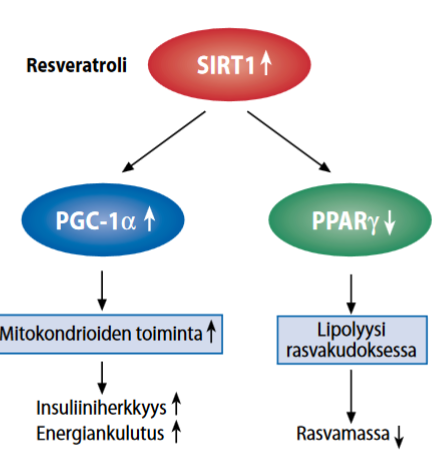
Insuliini ja terveys: Hiilihydraatti-insuliinimalli
Käsittelin edellisessä artikkelissa (Insuliini ja terveys: Johdanto) insuliinin aineenvaihduntaa ja toimintamekanismeja. Insuliini liittyy moniin elintapasairauksiin, kuten lihavuuteen, jota tarkastelen tässä hiilihydraatti-insuliinimallin kautta.
Insuliinin säätelemä anabolinen aineevaihdunta rakentaa uutta kudosta, kuten lihaksia ja rasvakudosta varastoimalla energiaa ja säätelemällä rasva- ja proteiinisynteesiä.
Rasvan varastoiminen läskinä ärsyttää monia, mutta se on fysiologisesti perusteltua. Lähes kaikki eläimet, mukaan lukien ihmiset, varastoivat energiaa rasvakudokseen. Niin luonto toimii. Läski on luonnon tapa varmistaa, että energiaa on saatavilla myös silloin, kun ravintoa ei ole saatavilla.
Kesällä lihova karhu ei pidä läskiä rumana. Läskin turvin karhu voi nukkua talven yli ja synnyttää talvipesässä terveitä ja vilkkaita karhunpentuja.
Koska läski on arvokasta energiaa, karhun ei tarvitse poistua pesästä etsimään ravintoa. Yhdessä grammassa karhun rasvakudokseen keräämää läskiä on yli tuplasti enemmän akkuvirtaa kuin grammassa hiilihydratteja tai proteiineja.
Rasvavarastojen täyttäminen kesällä ja talviuni talvella on karhun keino sopeutua talven ravintopulaan. Läski on ihmisen elimistön tapa turvata energian saanti. Lihominen on evoluution kannalta perusteltua.
Karhun lihomiseen kiteytyy läskin filosofia

Miten insuliini vaikuttaa lihomiseen?
Lihominen muuttuu herkästi sairaalloiseksi. Se vaikuttaa terveyteen ja sosiaaliseen elämään.
Tri Fatima Cody Stanford käyttää lihavuudesta samanlaista terminologiaa, kuin muista sairauksista: ihmisellä voi olla diabetes, tai hän voi sairastaa lihavuutta. Hän siis haluaa poistaa lihavuuteen liittyvät laiskuuteen ja ylensyöntin viittavat stigmat. Lihavuus on elämäntapasairaus.
Lihavuus on usein seuraus aineenvaihdunnan häiriintymisestä aivan kuten diabetes.
Hormonit säätelevät aineenvaihduntaa ja kertovat aivoille ravinnontarpeesta. Jos ja kun tämä järjestelmä menee sekaisin, ihminen alkaa varastoida energiaa sen sijaan, että kuluttaisi sitä. Tähän vaikuttaa erityisesti insuliini.
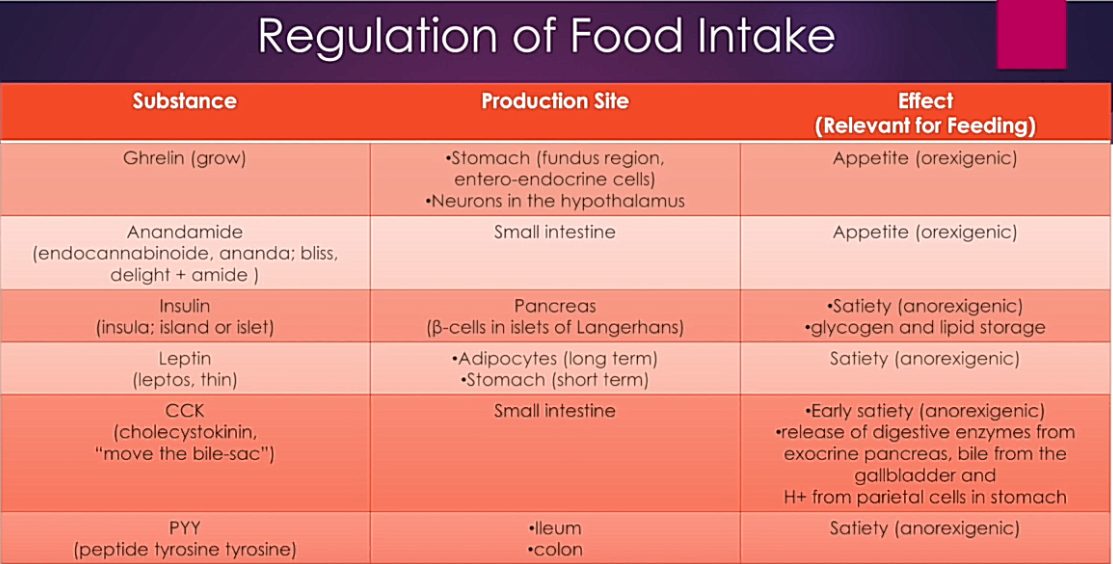
Rasvakudoksella on tärkeä fysiologinen rooli. Sen lisäksi, että rasvakudos varastoi energiaa ja turvaa energiansaannin silloin, kun ravintoa ei ole tarjolla, rasvakudos säätelee aktiivisesti kylläisyyden tunnetta leptiini-hormonin välityksellä. Nälän tunnetta säätelee suolistosta erittyvä greliini.
Biologiaa ja kemiaa ei oikein voi paeta. Informaatioajan bittivirtojen some-olentoina olemme kuitenkin hormonien välittämiä signaaleja tyydyttäviä eläimiä.
Hormonien vaikutuksen ymmärtäminen voi auttaa ymmärtämään nälkää ja lihottavia mielihaluja. Hormonitoiminnan häiriintyminen auttaa ymmärtämään lihavuutta aineenvaihdunnan sairautena.
Rasvasotaa
Ihminen lihoo syömällä liikaa. Se ei ole mikään salaisuus. Todellinen mysteeri on se, miksi ihminen syö liikaa ja lihoo silloinkin, kun hänellä on rasvavarastoissaan kuukaudeksi energiaa.
Lihavien määrä on kolminkertaistunut vuoden 1980 jälkeen. Intensiivisestä tutkimuksesta huolimatta globaalin lihavuusepidemian syyt ovat yhä arvailujen ja kiistojen aihe.
Rasvasota ei päättynyt 1970-luvulla, vaan jatkuu siirappisena asemasotana. Tämän sodan sokereiden sairastuttamat siviiliuhrit kamppailevat nyt lihavuuden, metabolisen oireyhtymän, rasvamaksan ja aikuisyypin diabeteksen lisäksi muisti- ja suolistosairauksia vastaan ahmimalla samaa korkean glykeemisen kuorman sokeripitoista roskaa, joka sai heidät alun alkaenkin sairastumaan. John Yudkin ja monet muut varoittivat tästä jo 1970-luvulla.
Aineenvaihduntaan liittyvät sairaudet kuormittavat taloutta ja kasvattavat terveydenhoitomenoja. Yhdysvalloissa lihavia on pian yhtä paljon kuin television omistajia.
Perinteisillä kalorirajoitetuilla dieeteillä ei ole pitkäkestoista laihduttavaa vaikutusta. Lihavuustutkimukset osoittavat, että vain yksi kuudesta kaloreita rajoittamalla laihtunut onnistuu välttämään yli 10 prosentin lihomisen laihduttamista seuraavan vuoden aikana. Lähes kaikki kaloreiden rajoittamiseen perustuvat laihdutuskuurit epäonnistuvat.

Dieettien ympärille on kasvanut hedelmällisiä laihdutus- ja hyvinvointibisneksiä, joiden rahavirtoja ohjaa usko perinteiseen kaloriteoriaan.
Ajatus, että kaikki kalorit ovat samanarvoisia, on hieman harhainen. Kalorien rajoittaminen johtaa usein painon jojoiluun, koska se hidastaa perusaineenvaihduntaa.
Miksi lihavalla on nälkä, vaikka hän karhuemon tapaan olisi varastoinut rasvakudokseen riittävästi energiaa koko talveksi?
Nälkää ja kylläisyyttä säätelee hormonitoiminta (erityisesti greliini ja leptiini). Insuliinin ja glukagonin eritystä säätelee ruokaan liittyvät aistimukset ja verensokeripitoisuuden muutokset.
Hormonaalisesti ravinnon kalorimäärää tärkeämpi tekijä on ravinnon laatu. Laadullisilla valinnoilla voi vaikuttaa nälän tunteeseen ja ohjata elimistö rasvan aktiiviseen polttamiseen betaoksidaatiossa. Se on oikeastaan aika helppoa.
CIM (hiilihydraatti-insuliinimalli)
Lihomisen hiilihydraatti-insuliinimallin (CIM/Carbohydrate-Insulin-Model) mukaan pitkälle jalostettujen, korkean glykeemisen kuorman ruokien kulutuksen kasvu määrittelee syömistavoissa tapahtunutta muutosta ja korreloi lihavuusepidemian kanssa.
Vuoden 1980 jälkeen yhä suurempi osa päivittäisestä energiasta on saatu sokereista (hiilihydraateista). Rasvan ja aivan erityisesti tyydyttyneen rasvan kulutus on samaan aikaan laskenut. Hiilihydraatit eivät pidä nälkää loitolla läheskään niin hyvin, kuin rasva, ja siksi ihmiset puputtavat sokereita aamusta iltaan.
Jatkuvasti korkea verensokeri ja verenkierrossa raivoava insuliinimyrsky vaikuttavat nälkää ja kylläisyyttä säätelevien hormonien lisäksi terveyteen.
Korkea verensokeri ja insuliini ovat tunnettuja verisuonia ja elimiä vaurioittavia sydän- ja verisuonitautien riskitekijöitä. Nykyinen hiilihydraattipainotteinen elämäntapa altistaa insuliiniresistenssille, lihomiselle, aikuistyypin diabetekselle, sydän- ja verisuonitaudeille, suolistosairauksille ja muistisairauksile.
Tämä kaikki on todistettu lukemattomissa in vivo ja in vitro ihmis- ja eläinkokeissa sekä laboratorioiden solumaljoissa.
Kuinka korkean glykeemisen kuorman ruoka vaikuttaa?
Hiilihydraattipainotteinen ruokavalio aiheuttaa hormonaalisia muutoksia, jotka vaikuttavat aineenvaihduntaan. Ne edistävät energian varastoimista rasvakudokseen, kasvattavat nälkää ja vähentävät energiankulutusta.
Aineenvaihdunnan toimintaa ja terveyttä ei paranna se, että hiilihydraatit ja suositut siemenöljyt ylläpitävät inflammaatiota tuottamalla vapaita happiradikaaleja. Liiallinen omega6-rasvahappojen saanti johtaa arakidonihapon muodostumiseen, josta elimistö syntetisoi prostaglandiineja. Prostaglandiinien synteesi elimistössä kiihtyy tavallisesti kuumeen ja kivun seurauksena, sillä ne vaikuttavat tulehdusreaktion syntyyn.

Tutkimuksista
Tutkimustulokset tukevat hiilihydraatti-insuliinimallia. Eläinten ruokavalio vaikuttaa CIM-mallin antaman ennusteen mukaisesti eläimen aineenvaihduntaan ja kehon koostumukseen kalorien määristä riippumatta.

Rottakoe: Samat makrot ja kalorimäärä. 3 viikon koe. Ainoana muuttujana omega6, jota lihakarjan talissa on vähän, oliiviöljyssä hieman enemmän saffloriöljyssä on valtavasti.

Omegakutosten vaikutus painoon ja kehon rasvakoostumukseen.
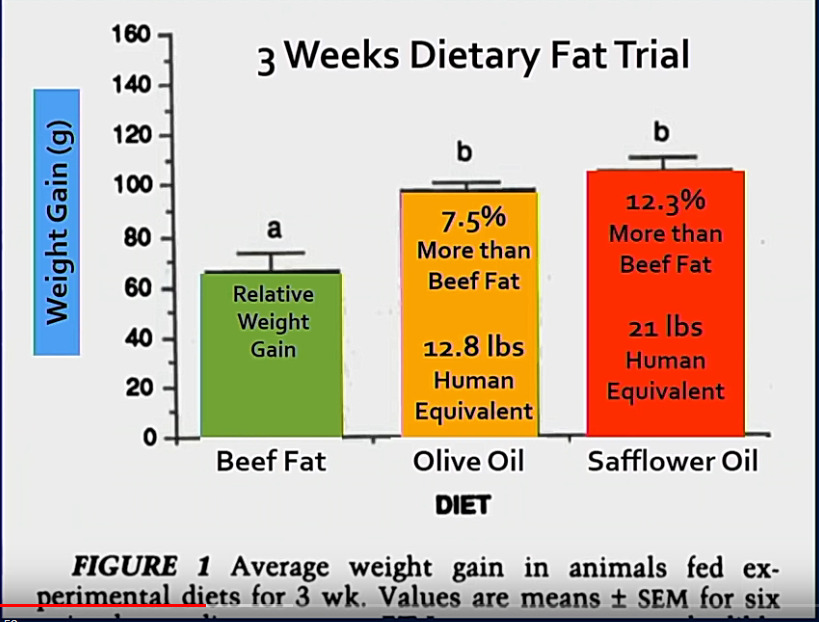
Kalorimäärät, eli rottien energiansaanti oli täsmälleen sama ja omegakutosia paitsi myös ravinnonlähteet olivat samoja. Näin suuri vaikutus painoon oli pelkästään omegakutosilla.
Käyttäytymistutkimusten meta-analyysit kertovat tehokkaammasta painonpudotuksesta vähäisellä glykeemisellä kuormalla verrattuna vähärasvaiseen ruokavalioon.
Verensokerin laskeminen ja insuliinitasojen pitäminen matalina vähän hiilihydraatteja ja runsaasti rasvaa sisältävällä ruokavaliolla laihduttaa varmemmin ja terveellisemmin kuin kaloreiden rajoittaminen. Se on terveellisin tapa hoitaa alkoholista riippumatonta rasvamaksaa, metabolista oireyhtymää ja aikuistyypin diabetesta.
Lääkeresistenttiä epilepsiaa on hoidettu jo sata vuotta hiilihydraatteja rajoittamalla. Nyt positiivisia hoitotuloksia saadaan myös Parkinsonin taudin ja Alzheimerin taudin hoidossa.
Pienen glykeemisen kuorman ravinto ja ketoosi kiinnostavat NASA:a ja Yhdysvaltojen puolustusministeriötä. Äärioloissa ketoosi on osoittautunut ylivoimaiseksi aineenvaihduntamekanismiksi. Siksi ketoosia tutkitaan taistelusukeltajien ja astronauttien toimintakyvyn tehostajana.
Vielä vuosikymmen sitten luultiin, että aivosolut tarvitsevat välttämättä glukoosia, mutta nyt tiedetään, että betahydroksibutyraatti on aivosoluille optimaalista ravintoa. Elimistö valmistaa betahydroksibutyraattia rasvasta, kun sokerin saanti loppuu. Neurodegeneratiivisia tauteja sairastavien koehenkilöiden aivojen magneettikuvissa sairauden hiljentämät alueet ovat aktivoituneet uudestaan ja kognitiiviset kyvyt parantuneet rasvapainotteisella ravinnolla. Erityisesti keskipitkät rasvahapot (MCT) ovat tutkimuksissa kohentaneet koehenkilöiden aivojen aktiivisuutta ja kognitiota.

Olemassaolevat tutkimukset vahvistavat kuitenkin matalan glykeemisen kuorman ruokien edistävän laihtumista ja terveyttä korkean glykeemisen kuorman ruokia paremmin.
Hiilihydraattien laadun ja määrän lisäksi hiilihydraatti-insuliinimallia voi hyödyntää käsitteellisenä kehyksenä, joka auttaa ymmärtämään, kuinka eräät ravintoaineet altistavat aineenvaihdunnan lihomista edistäville hormonaalisille muutoksille.
Korkean glykeemisen kuorman ruokien rajoittaminen on käytännöllinen laihdutusmenetelmä, jonka painopiste ei ole rasvan ja kaloreiden vähentämisessä, vaan verensokeri- ja insuliinitasojen madaltamisessa.
Syö vähemmän” ja ”liiku enemmän!
Me opimme tuon imperatiivin jo peruskoulussa. Voipaketti, pekoni ja kananmunat palauttavat mieleen vanhat pelot: yydyttynyt rasva on lihottavaa myrkkyä, joka murhaa sinut hitaasti, mutta varmasti tukkimalla verisuonesi.
Pelottavien kummitusjuttujen rinnalla olemme oppineet, että siemenöljyt ja margariinit ovat superhyperextraterveellisiä rasvoja, jotka laskevat vastenmielistä kolesterolia. Se on pitkälti roskaa!
Tieteellinen näyttö, ihmisen fysiologia ja miljoonien ihmisten henkilökohtaiset kokemukset osoittavat, että painonpudotuksen myötä nälkä lisääntyy ja energiankulutus laskee.
Toisin sanoen laihduttajan paino jumittaa nälkäkuurilla ja nälkä kurnii vatsanpohjassa. Useimmat kaloreita rajoittamalla laihduttavat lihovat lähtöpainoon pian laihdutuskuurin jälkeen. Tiukka kaloreiden rajoittaminen hidastaa perusaineenvaihduntaa, minkä vuoksi ihminen lihoo entistä helpommin. Se voi olla ongelma laihduttajalle, mutta ei laihdutusbisnekselle.
Hiilihydraattien rajoittaminen ei edellytä kallista kuntosalikorttia tai hintavia laihdutusruokia. Sen sijaan vähähiilihydraattinen, runsaasti rasvaa ja kohtuullisesti proteiinia sisältävä ruokavalio pudottaa tehokkaasti painoa, tehostaa rasvan polttoa, laskee verensokeria ja verenpainetta.
Rasvapainotteinen ruokavalio voi nostaa lipoproteiinien määrää (kolesterolia), mutta elimistö säätelee kolesterolin homeostaasia. Lipoproteiinien määrä lisääntyy, koska LDL ja HDL ovat rasvaa, kolesterolia ja rasvaliukoisia vitamiineja soluihin kuljettavia molekyylejä.
Kun solujen energia otetaan rasvasta, veressä kulkee luonnollisesti enemmän rasvaa kuljettavia molekyylejä
Lisääntyvä tutkimusnäyttö rauhoittelee kolesteroliin ja tyydyttyneisiin rasvoihin liittyviä pelkoja. Kolesteroli on elintärkeä aine ja tyydyttyneet rasvat ovat sydäntautien osalta pahimmillaankin neutraaleja.
On siis aika kääntää uusi lehti terveystiedoissa: sydän- ja verisuonitaudit johtuvat kasvavan näytön perusteella jatkuvasti korkeasta verensokerista ja insuliinista. Kolesteroli on todennäköisesti seuraus verisuonten vahingoittumisesta, ei syy vahintoittumiselle.
Ajatus laihdutuskuurista on muutenkin nurinkurinen. Teoria, että laihduttaja vähentää energian saantia, kunnes tavoitepaino saavutetaan ja palaa ihannepainoisena ruokavalioon, joka johti lihomiseen, on yksinkertaisesti kestämätön.
Pysyvä laihtuminen edellyttää elämäntaparemonttia. Pelkkä laihdutuskuuri ei riitä, koska paino palaa korkojen kera heti kun kuuri loppuu.
Matalan glykeemisen kuorman ruokavalio (esimerkiksi ketogeeninen ruokavalio) laskee painoa käyttämällä rasvakudokseen varastoitua energiaa.
Hiilihydraattipainotteinen ruokavalio estää rasvan käyttämisen energianlähteenä, koska insuliini on lipolyysin estäjä. Kaloreita rajoittamalla laihduttaja kannibalisoi lihaksiaan vähintään yhtä paljon kuin rasvakudosta. Ketogeeninen aineenvaihduntaan perustuva ruokavalio käynnistää lipolyysin, jolloin keho saa tarvitsemansa energian rasvakudoksesta, eikä elimistön tarvitse pilkkoa lihaksia ravinnoksi. Tämä on osoitettu mm. Minnesota Starvation Experimentissä 1944-1945.

Hiilihydraatti-insuliinimalli
Vaihtoehtoisen näkemyksen mukaan syömistottumusten ja syödyn ravinnon muutokset johtavat hormonaalisiin reaktioihin, jotka ohjaavat ravinnosta saatua energiaa rasvasoluihin.
Hiilihydraatti-insuliinimallissa rasvasolujen rooli on aktiivinen lihavuuden etiologian kannalta. Rasvakudos kertoo leptiinin välityksellä aivoille kehon energiabalanssista. Tämän mekanismin häiriintyminen, leptiiniresistenssi, johtaa tilaan, jolloin ihminen kokee itsensä nälkäiseksi, vaikka hän olisi hetkeä aimmin täyttänyt itsensä runsasenergisella aterialla.
Vaikka monet tekijät vaikuttavat rasvan kumuloitumiseen, anabolisena hormonina insuliini heiluttelee tahtipuikkoa energia-aineenvaihdunnan säätelyssä ja ravinteiden varastoimisessa.
Insuliini vähentää kaikkien energiaravinteiden kiertokonsentraatiota stimuloimalla glukoosinottoa kudoksiin, tukahduttamalla rasvahappojen vapautumista rasvakudoksesta, estämällä ketonien tuotantoa maksassa ja edistämällä rasvan ja glykogeenin varastoimista.
Insuliini on lipolyysin estäjä
Insuliini on kolmen lipolyysiin osallistuvan entsyymin estäjä. Estämällä hormonisensitiivistä lipaasia (HSL), rasvakudoksen triglyseridilipaasia (ATGL) ja epäsuorasti CPT1-entsyymiä, insuliini estää rasvasolujen purkamisen vapaiksi rasvahapoiksi lipolyysissä.
Veren kohonnut insuliinipitoisuus (hyperinsulinemia) esimerkiksi tyypin 2 diabeteksen insuliinihoitojen aloittamisen jälkeen ennustaa suurella todennäköisyydellä painonnousua.
Osa insuliinin aiheuttamasta painonnoususta liittyy aineenvaihdunnan muutoksiin. Tyypin 1 diabeteksen haiman Langerhansin saarekkeiden beetasolujen tuhoutuminen, riittämätön insuliinihoito tai insuliinin eritystä estävät lääkkeet aiheuttavat tunnetusti laihtumista. Insuliinin vaikutukset painoon on kattavasti dokumentoitu.
Haima on erityisen herkkä hiilihydraateille. Verensokerin kohoaminen johtaa moninkertaiseen insuliinin eritykseen muihin ravintoaineisiin verrattuna. Glukoosipitoisuuden kohoaminen veressä johtaa insuliinivasteeseen, joka on jotakuinkin kymmenkertainen rasvan aiheuttamaan insuliinivasteeseen nähden. Myös hiilihydraattien määrällä ja laadulla on selvä vaikutus insuliinin eritykseen.

Glykeeminen kuorma
Glykeeminen indeksi (GI) kuvaa, kuinka nopeasti tietyt elintarvikkeet nostavat verensokeria (ja siten insuliinia) 2 tunnissa syömisen jälkeen.
Suurin osa valkoisista jauhoista, perunaruoista ja lisätyistä sokereista sulavat nopeasti ja niillä on korkea GI, kun taas ei-tärkkelystä sisältävillä vihanneksilla, palkokasveilla, kokonaisilla hedelmillä ja täysjyvillä on yleensä matalampi GI.
Tähän liittyvä mitta, glykeeminen kuorma (GL, Glycemic Load) on paras ruokailun jälkeisen verensokerin ennustaja. Se selittää jopa 90% varianssista. Glykeeminen kuorma kertoo ruoan sisältämien hiilihydraattien laadusta ja määrästä.
Tämä on tuttua tyypin 1 diabetesta sairastaville, mutta monet tyypin 2 diabetekseen sairastuneet jatkavat lääkkeiden avustuksella lihottavan ja sairautta ylläpitävän ruokavalion noudattamista.
Tyypin 2 diabeteksen saa remissioon hiilihydraatteja rajoittamalla, esimerkiksi ketogeenisellä ruokavaliolla. Tätä hoitomuotoa suosittelee jo yli 10 000 lääkäriä ympäri maailman. Se on tehokkain tapa hoitaa tyypin 2 diabetesta, alkoholista riippumatonta rasvamaksaa ja metabolista oireyhtymää.
Glykemiakuormaa voidaan hyödyntää arvioitaessa aterian vaikutusta veren sokeriin ja veren insuliinivasteeseen. Vaikka aterialla nautittaisiin ruoka-aineita, joilla on korkea glykeeminen indeksi (GI), ei vaikutus veren sokeriin ole kovin suuri jos näitä ruoka-aineita on vain vähän suhteessa aterian kokoon. Tällöin aterian glykemiakuorma on pieni. Suuri hiilihydraattimäärä ja glykeeminen indeksi taas kasvattavat myös glykemiakuormaa.
Glykemiakuorma lasketaan seuraavasti: GI x imeytyvän hiilihydraatin määrä / 100. Aterian glykemiakuormaa määritettäessä lasketaan yhteen sen sisältämien ruoka-aineiden GL-arvot.
 Proteiini stimuloi insuliinin eritystä aminohappokoostumuksesta riippuen, mutta proteiini vaikuttaa myös insuliinin vastavaikuttajan, katabolisen glukagonin, eritykseen. Ravinnon rasvalla on vain vähän suoraa vaikutusta insuliinin eritykseen, mikä muodostaa teoreettisen perustan runsaasti rasvaa sisältävien ruokavalioiden terveys- ja laihdutusvaikutuksille.
Proteiini stimuloi insuliinin eritystä aminohappokoostumuksesta riippuen, mutta proteiini vaikuttaa myös insuliinin vastavaikuttajan, katabolisen glukagonin, eritykseen. Ravinnon rasvalla on vain vähän suoraa vaikutusta insuliinin eritykseen, mikä muodostaa teoreettisen perustan runsaasti rasvaa sisältävien ruokavalioiden terveys- ja laihdutusvaikutuksille.
Hiilihydraatti-insuliinimallin mukaan runsashiilihydraattinen, erityisesti lisättyä sokeria ja/tai tärkkelystä sisältävä ruokavalio aiheuttaa aterianjälkeistä hyperinsulinemiaa, mikä lisää energian varastoimista rasvasoluihin altistaen lihomiselle.
Ylimääräinen glukoosi varastoidaan rasvasoluihin, koska veressä glukoosi voi aiheuttaa glykaation muiden ravinteiden kanssa.
Glykaatio
Glykaatio on mikä tahansa ei-entsymaattinen reaktio, jossa glukoosi tai mikä tahansa muu hiilihydraatti liittyy kovalenttisesti muunlaisiin molekyyleihin kuten proteiineihin, lipideihin tai DNA:han. Kehittynyt glykaation lopputuote (eng. advanced glycation end-product, AGE) on yleisnimitys suurelle joukolle erilaisia proteiineja tai rasvoja, jotka ovat glykatoituneet ei-entsymaattisesti altistuttuaan joillekin sokereille, eli liittyneet näihin sokereihin.AGE:ja voidaan myös kutsua glykotoksiineiksi ja ne saattavat olla pahentava osatekijä ikääntymisessä ja osallistua useiden rappeuttavien sairauksien kuten Alzheimerin taudin, diabeteksen, valtimonkovettumataudin ja kroonisen munuaisten vajaatoiminnan kehittymiseen. Lisäksi niiden on tutkittu osallistuvan harmaakaihin, lihasheikkouden ja syöpäkasvainten kehittymiseen.

CIM vs. kalorimalli
Kuten perinteinen kalorimalli, myös CIM pohjautuu termodynamiikan ensimmäiseen lakiin, jossa verrataan energiansaannin ja energiankulutuksen erotusta.
CIM pitää ylensyöntiä rasvan määrän lisääntymisen seurauksena, ei rasvan määrän ensisijaisena syynä.
Toisin sanoen hiilihydraatti-insuliinimallissa syy-seuraus-suhde, joka yhdistää energian tasapainon rasvakudokseen, virtaa vastakkaiseen suuntaan hormonaalisten signaalien ohjaamana. Tästä näkökulmasta kalorien rajoittamista voidaan pitää oireenmukaisena hoitona, joka valitettavan usein epäonnistuu nykyaikaisessa ravintoympäristössä.
Vähäkaloriset / vähärasvaiset ruokavaliot voivat pahentaa taustalla olevaa aineenvaihdunnan häiriötä rajoittamalla edelleen veressä olevan energian hyödyntämistä ja käynnistämällä nälkävasteen, johon liittyy kasvanut nälän tunne, aineenvaihdunnan hidastuminen ja kohonnneet stressihormonitasot.
Eläintutkimusten kertomaa
Insuliinin injektio keskushermostoon aiheuttaa ruokahaluttomuutta ja laihtumista, mutta insuliinin antaminen perifeerisesti edistää rasvan varastoitumista rasvakudokseen, kasvattaa nälän tunnetta ja aiheuttaa painonnousua.
Vaikka koe-eläinten kaloreita rajoitetaan painonnousun estämiseksi, insuliinia saaneilla eläimilä rasvakudokseen varastoituu yhä rasvaa, mikä vastaa CIM:n ennustetta energiatalouden muuttuneista painopisteistä.
Ruokavalioilla, jotka luontaisesti lisäävät insuliinin eritystä, on samanlaisia metabolisia vaikutuksia kuin insuliinin injektiolla.
Jyrsijöillä, joita ruokitaan korkean glykeemisen kuorman ruokavaliolla, ilmenee progressiivisia poikkeamia aineenvaihdunnasa. Näitä ovat:
- hyperinsulinemia
- rasvasolujen koon kasvu
- lisääntynyt rasvan varastointi
- pienempi energiankulutus
- kasvanut nälkä
Kaloreiden rajoittaminen painonnousun estämiseksi eläimillä, jotka saivat korkean glykeemisen kuorman ravintoa, ei estänyt lihomista tai siihen liittyviä kardiometabolisia riskitekijöitä. Koe tuki insuliininantokokeiden tuloksia.
Sen sijaan jyrsijöiden energiankulutus kasvoi ja paino laski niillä hiirillä, jotka saivat hyvin vähän hiilihydraatteja verrattuna tavanomaisella ruokavaliolla ruokittuihin hiiriin. Tämä toteutui, vaikka hiirien ravinnon kalorimääräisessä saannissa ei ollut eroja.

Geneettiset mallit
Korkeat insuliinipitoisuudet veressä voivat johtua primaarisesta insuliinin ylierityksestä (joka usein aiheuttaa painonnousua) tai olla vaste insuliiniresistenssille (joka voi suojata painonnousulta, etenkin jos insuliiniresistenssiä esiintyy rasvakudoksessa. Siksi paastoinsuliiniin ja painoon perustuvat havainnointitutkimukset eivät anna luotettavaa CIM-ennustetta.
Geneettisten tutkimusten avulla voidaan havaita syy-seuraus-suhteita
”In a recent report, bi-directional Mendelian Randomization was used to examine the relationship between insulin secretion and BMI, potentially free from confounding by socio-demographic and behavioral factors inherent to most conventional associational analyses. This study found that genetically-determined insulin secretion strongly predicted BMI , whereas genetically-determined BMI did not predict insulin secretion. In addition, variants in the insulin promotor gene associated with insulin hypersecretion in humans predict weight gain during adolescence. Furthermore, transgenic mice with reduced insulin secretion had increased energy expenditure and were protected from diet-induced obesity, leading the investigators to conclude, in accordance with the CIM, that “circulating hyperinsulinemia drives diet-induced obesity and its complications.”
Käyttäytymiskokeet ja havainnointitutkimukset
Toisin kuin tavanomaisessa lihomismallissa ennustetaan, vähärasvaisten ruokavalioiden luonnostaan pienempi energiamäärä ei itsestään ylläpidä jatkuvaa laihtumista.
Useissa viimeaikaisissa meta-analyyseissä on osoitettu, että vähärasvaiset ruokavaliot ovat pitkällä aikavälillä huonompia laihdutusruokavalioita kuin korkeamman rasvapitoisuuden (vähähiilihydraattiset) ruokavaliot.
”Of note, two major trials that employed special measures to improve compliance, Diogenes and the DIRECT trial found greater weight loss on low- vs high-GL diets A third major study, DIETFITS, reported non-significantly more weight loss on a Healthy Low-Carbohydrate Diet vs Healthy Low-Fat Diet, but both groups were counselled to avoid refined grains, sugar and other processed foods. Consequently, the GL of the Healthy Low-Fat Diet was exceptionally low for a higher-carbohydrate diet – similar to that of the lowest-GL diet in Diogenes.”
Suurissa, pitkäkestoisissa kohorttitutkimuksissa eräillä erittäin rasvaisilla elintarvikkeilla (esim. pähkinät ja täysrasvaiset maitotuotteet), on joko olematon tai käänteinen yhteys painonnousuun. S
itä vastoin monet runsaasti kulutetut korkean glykeemisen kuorman elintarvikkeet (esim. perunat, riisi, maissi, vaaleat viljat, makeat jälkiruoat, sipsit, sokerijuomat ja hedelmämehut) assosioituivat vahvasti lihomiseen.

Ravintotutkimukset
Hiilihydraatti-insuliinimallin mukaan korkean glykeemisen kuorman ateria rajoittaa energiaravinteiden saantia noin 3-5 tuntia syömisen jälkeen, vähentää rasvan polttoa, pienentää energiankulutusta, stimuloi stressihormonin eritystä ja kasvattaa nälkää.
Nämä vaikutukset on raportoitu useissa tutkimuksissa. Pitkällä aikavälillä rasvan varastointi lisääntyy korkean glykeemisen kuorman ruokavalion noudattamisen seurauksena.
Tavanomaisessa runsaasti hiilihydraatteja sisältävässä ruokavaliossa aivot ovat riippuvaisia glukoosista (yli 100 g / päivä). Ketogeenisen ruokavalion alkuvaiheessa aineenvaihdunnan on tuotettava glukoosia verenkierron vapaista aminohapoista, sitruunahappokierron väliaineista ja glyserolista maksassa tapahtuvassa glukoneogeneesissä.
Jos veressä ei ole riittävästi vapaita aminohappoja glukoneogeneesiin, kehon on väliaikaisesti katabolisoitava (tai kannibalisoitavat) lihasten proteiineja glukoneogeneesin lähtöaineiksi. Tämä on kuitenkin väliaikainen vaihe, koska ketogeneesin tuottamien ketonien pitoisuus ja merkitys aivojen ravintona lisääntyy muutamassa päivässä.
Hyvin vähähiilihydraattisen ruokavalion (ja pitkittyneen paaston) tunnusmerkki on ketoosin kehittyminen – josta on peräisin nimitys ”ketogeeninen ruokavalio”.
Nälkää käsittelevät tutkimukset kertovat rasva-aineenvaihduntaan sopeutumisen etenemisestä.
Owen et al., osoitti, että ketonipitoisuus: ß-hydroksibutyraatti, asetoetikkahappo ja asetoni lisääntyivät asteittain 10 päivän ajan ja saavuttavat vakaan tason vasta noin 3 viikon paaston jälkeen.
Yang et al. osoitti, että ketonien erittyminen virtsaan lisääntyi 10 päivän ajan hyvin vähähiilihydraattisella ruokavaliolla, mutta hitaammin kuin paaston aikana.
Vazquez et al. osoitti, että typpitasapaino oli negatiivisempi vähäkalorisella ketogeenisellä ruokavaliolla verrattuna ei-ketogeeniseen ruokavalioon noin 3 viikon ajan, ja saavutti sitten neutraalin nettotasapainon (eli ei lisännyt rasvattoman massan menetystä).
On hyvin dokumentoitu, että rasva-aineenvaihduntaan sopeutuminen edellyttää vähintään 2-3 viikon vähähiilihydraattisen ruokavalion noudattamista. Ehkä pidempäänkin. Lyhyemmät tutkimukset eivät osoita makroravintoaineiden aiheuttamia aineenvaihdunnan muutoksia.

Liiallinen syöminen aiheuttaa liikalihavuutta
Kalorien liiallinen kulutus johtaa painonnousuun, kuten termodynamiikan ensimmäisessä laissa määritellään. Keho pyrkii kuitenkinsäilyttämään painon tasapainossa. Pitkällä aikavälillä liiallinen energian saanti kasvattaa dynaamisesti energiankulutusta ja vähentää nälän tunnetta. Nämä fysiologiset mekanismit pyrkivät vähentämään jatkuvaa painonnousua.
Klassisissa yliruokintatutkimuksissa monet kokeisiin osallistuneet ilmoittivat tuntevansa olonsa epämukavaksi liiallisella energian saannilla. Useilla oli vaikeuksia jatkaa liian paljon energiaa sisältävää ruokavaliota. Kun ohjelmat päättyivät, useimpien kokeisiin osallistuneiden ruumiinpaino palasi spontaanisti lähtötasolle tai sen lähelle.
Eläimillä ja ihmisillä tehdyt tutkimukset vahvistavat, että fysiologiset tekijät rajoittavat liiallista lihomista ja liiallista painonlaskua. Näiden mekanismien häiriintyminen (sairastuminen) aiheuttaa lihomista.
Hiilihydraatti-insuliinimallin mukaan korkean glykeemisen kuorman ruokavalio muuttaa näitä luonnollisia homeostaattisia mekanismeja epäterveelliseen suuntaan.
Lihominen liittyy tyypillisesti verenkierron kohonneisiin glukoosi-, insuliini- ja rasvahappopitoisuuksiin
”Unfortunately, cross-sectional studies after development of obesity may also confound understanding of etiology. The CIM proposes that metabolic fuel concentration is reduced with a high-GL diet in the late postprandial period (approximately 2.5 to 5 hr after eating) due to excessive adipose anabolic activity during the dynamic stage of obesity development. Eventually, fat cells reach a limit, beyond which they cannot effectively expand storage capacity.At this stage, weight gain plateaus (at the cost of increasing insulin resistance and chronic inflammation) and circulating metabolic fuel concentrations consequently rise.”
Toisaalta Yhdysvalloissa proteiinin ja rasvan absoluuttinen saanti ei ole juurikaan muuttunut 1970-luvulta lähtien, kun taas hiilihydraattien, (pääasiassa korkean glykeemisen kuorman valkoisten jauhojen, perunoiden, pikakruoan ja lisätyn sokerin) saanti on lisääntynyt huomattavasti, mikä on kasvattanut merkittävästi kalorien kokonaissaantia.
Kansainväliset epidemiologiset tutkimukset eivät aina osoita selkeää rinnakkaisuutta korkean glykeemisen kuorman ja liikalihavuuden esiintyvyyden välillä.
Historiallisesti maatalousyhteisöjen aasialaiset pysyivät hoikkina runsashiilihydraattisella riisipohjaisella ruokavaliolla.Tätä voi selittää aasialaisten luonnollisesti vähäisempi rasvakudoksen määrä, niukka ravinnon saanti ja korkea energiankulutus.
Japanissa lihavia on nelisen prosenttia väestöstä, vaikka diabetesta sairastavia on yli seitsemän prosenttia. Lihavuus ja aikuistyypin diabetes kulkevat usein käsikkäin. Yhdysvalloissa lihavia on lähes kolmannes väestöstä ja diabetesta sairastavia yksi kymmenestä.
Kun fyysisen aktiivisuuden taso on laskenut kaupungistumisen myötä (esim. Kiina), lihavien ja diabetesta sairastavien määrät ovat nousseet dramaattisesti.
Kliiniset vaikutukset
CIM on käytännöllinen vaihtoehto lihavuuden, metabolisen oireyhtymän ja diabeteksen hoitoon. Viime aikoina on osoitettu, että tavanomaiset vähärasvaiset ja vähäkaloriset ruokavaliot eivät hillitse lihavuus- ja diabetesepidemioita.
Ensisijainen painopiste olisi asetettava kulutettujen kaloreiden laatuun eikä määrään, jotta kalorien jakaminen siirrettäisiin pois rasvakudoksen varastoinnista ja parannettaisiin energiaravinteiden saatavuutta muuhun kehoon. Tällä tavoin negatiivinen energiatasapaino ja laihtuminen voidaan saavuttaa helpommin ja kestävämmillä tuloksilla.
Ruokavalioilla, joilla on erilainen makroravinnekoostumus, on kaloripitoisuuden erojen lisäksi erilaisia vaikutuksia hormoneihin, aineenvaihduntareiteihin, geenien ilmentymiseen ja suolen mikrobiomeihin. Nämä vaikuttavat rasvan varastointiin.
Väittämällä, että kaikki kalorit ovat samanarvoisia keholle, perinteinen malli sulkee pois ympäristötekijät, jolla on kaikkein vahvin yhteys painonhallintaan. Mitkä muut tekijät voisivat selittää lihavuuden esiintyvyyden kasvua 1980-luvulta, kuin samaan aikaan tapahtuneet merkittävät muutokset syömistavoissa?
Hiilihydraatti-insuliinimalliin perustuvat ruokavaliosuositukset
- Vähennä valkoisia viljoja, perunatuotteita ja lisättyjä sokereita sekä korkean glykeemisen kuorman hiilihydraatteja, joiden yleinen ravitsemuksellinen laatu on heikko
- Suosi matalan glykeemisen kuorman hiilihydraatteja, mukaan lukien ei-tärkkelyspitoiset vihannekset, palkokasvit ja ei-trooppiset kokonaiset hedelmät (kuten omenat)
- Jos syöt viljatuotteita, valitse kokojyväviljat ja perinteisesti jalostetut vaihtoehdot (esim. Kokonainen ohra, quinoa ja perinteiseen taikinajuureen tehty leipä
- Lisää pähkinöitä, siemeniä, avokadoa, oliiviöljyä ja muita terveellisiä rasvaisia ruokia
- Syö riittävästi proteiinia (myös kasviperäistä)
- Hiilihydraattien saannin rajoittaminen ja korvaaminen rasvalla tuottaa suurimman hyödyn
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6082688/
http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/pancreas/insulin_phys.html
https://www.verywellhealth.com/how-insulin-works-in-the-body-1087716
https://www.ncbi.nlm.nih.gov/books/NBK525983/
https://thefastingmethod.com/understanding-joseph-kraft-diabetes-in-situ-t2d-24/
https://denversdietdoctor.com/diabetes-vascular-disease-joseph-r-kraft-md/
https://dopinglinkki.fi/en/info-bank/doping-substances/insulin